L’hivern de la fam
Els bessons univitel·lins tenen exactament el mateix codi genètic l’un i l’altre. Comparteixen el mateix úter, i acostumen a criar-se en ambients molt semblants. Tenint això en compte, sembla lògic que, si un dels bessons contrau esquizofrènia, la probabilitat que el seu germà o la seva germana també la contregui sigui molt alta. Potser hauríem de preguntar-nos, fins i tot, per què no és més alta encara. ¿Per què no és del cent per cent?
NESSA CAREY, La revolució epigenètica
Al llarg del segle XX, els gens han seguit una trajectòria sensacional […]. Ens han conduït fins a les portes d’una nova era de la biologia, una era que conté la promesa d’avenços encara més espectaculars. Aquests mateixos avenços, però, exigiran la introducció d’altres conceptes, altres termes i altres maneres de pensar sobre l’organització biològica, que reduiran la influència que els gens han tingut en la concepció de les ciències de la vida.
EVELYN FOX KELLER, Una antropologia de la biomedicina
Al capítol anterior ha quedat una pregunta implícita per respondre. Si la «persona» es genera per interaccions fortuïtes entre els esdeveniments i els gens ¿com queden registrades a la pràctica aquestes interaccions? De dos germans bessons, un cau sobre el gel, es trenca un genoll i desenvolupa una callositat òssia, i l’altre bessó no. Una germana es casa amb un executiu de Delhi amb futur, i l’altra se’n va a viure en una casa tronada de Calcuta. ¿Per mitjà de quins mecanismes aquestes «accions del destí» queden registrades en una cèl·lula o un cos? La resposta ha sigut automàtica des de fa dècades: per mitjà dels gens; o, si volem ser més exactes, activant els gens o desactivant-los. A París, a la dècada del 1950, Monod i Jacob havien demostrat que, quan els bacteris passen de la dieta de glucosa a la de lactosa, reprimeixen els gens metabolitzadors de la glucosa i activen els gens metabolitzadors de la lactosa. (Aquests gens s’activen i es reprimeixen per mitjà de factors de regulació general —activadors i repressors— coneguts també amb el nom de factors de transcripció). Cap a trenta anys més tard, uns biòlegs que treballaven amb el cuc C. elegans havien descobert que els senyals de les cèl·lules veïnes —fets del destí, des del punt de vist d’una cèl·lula individual— també són detectats pels gens mestres activadors i repressors de regulació general, amb les consegüents alteracions en els llinatges cel·lulars. Quan un bessó cau al gel, els gens cicatritzadors s’activen. Aquests gens fan possible que la ferida s’endureixi fins a formar la callositat òssia al lloc de la fractura. Fins i tot quan al cervell es grava un record complex, els gens han de ser activats o reprimits. Quan un ocell canor descobreix un refilet nou d’un altre ocell, al cervell se li activa un gen anomenat ZENK. Si el refilet no és com hauria de ser —si és un refilet d’una altra espècie, o és desafinat—, el gen ZENK no s’activa amb la mateixa intensitat, i el refilet no s’emet.
Ara bé, aquesta activació o repressió dels gens a les cèl·lules i al cos (com a resposta a una successió d’efectes ambientals: caiguda, accident, cicatriu) ¿deixa alguna mena de senyal o d’empremta en el genoma? ¿Què passa quan un organisme es reprodueix? Els senyals o empremtes del genoma ¿es transmeten a un altre organisme? La informació sobre l’entorn ¿pot transmetre’s de generació en generació?
* * *
Estem a punt d’entrar en un dels terrenys més controvertits de la història del gen, i convé situar-lo en el seu context històric. A la dècada del 1950, l’embriòleg anglès Conrad Waddington va voler entendre els mecanismes per mitjà dels quals els senyals ambientals podien afectar el genoma d’una cèl·lula. En el desenvolupament embrionari, Waddington va poder observar la gènesi de milers de tipus cel·lulars diferents —neurones, cèl·lules musculars, sang, esperma— a partir d’una única cèl·lula fecundada. Recorrent a una analogia inspirada, Waddington va comparar la diferenciació embrionària amb una infinitat de bales de vidre rodolant per un paisatge en pendent ple de crestes, valls i esquerdes. Segons ell, quan cada cèl·lula recorria el seu camí particular de baixada per aquest «paisatge de Waddington», quedava atrapada en alguna canal o cresta concreta, cosa que determinava el tipus de cèl·lula en què s’acabaria convertint.
Waddington estava especialment intrigat per la forma en què l’entorn d’una cèl·lula podia afectar la utilització dels seus gens. Va percebre que aquest destí cel·lular havia d’estar determinat més enllà del genoma, i va batejar aquest fenomen amb el nom d’epi-genètica, que literalment significa ‘per sobre dels gens’.[069] L’epigenètica, segons Waddington va escriure, tracta de «la interacció dels gens amb l’ambient […] que fa que el fenotip aparegui».
* * *
Un macabre experiment humà va demostrar la teoria de Waddington, tot i que els seus resultats van trigar generacions a fer-se visibles. El setembre del 1944, enmig de la fase més carregada d’odi de la Segona Guerra Mundial, les tropes alemanyes que ocupaven Holanda van prohibir l’exportació d’aliments i de carbó a les regions septentrionals. Es va interrompre la circulació de trens i es van bloquejar les carreteres. Els canals, glaçats, no permetien la navegació. Van destruir amb explosius les grues, els vaixells i els molls del port de Rotterdam, cosa que va deixar, en paraules d’un locutor de ràdio, una «Holanda turmentada i exsangüe».
Holanda, tota ella solcada de canals i de barcasses, no només estava turmentada i exsangüe, sinó també afamada. Amsterdam, Rotterdam, Utrecht i Leiden depenien del transport regular per proveir-se d’aliments i de combustible. Al començament de l’hivern del 1944, el racionament que arribava a les províncies del nord dels rius Waal i Rin es va reduir a un degoteig, i la població estava a punt de patir una crisi de fam. El mes de desembre es van tornar a obrir els canals, però l’aigua ja s’havia glaçat. La primera cosa que va desaparèixer va ser la mantega, i a continuació el formatge, la carn, el pa i la verdura. La gent, morta de desesperació, de fred i de gana, arrencava els bulbs de les tulipes del jardí de casa seva i menjava les peles de les hortalisses, i més tard va passar a menjar escorça de bedoll, fulles i herba. Al final, la ingestió d’aliment va caure fins a unes quatre-centes calories diàries, l’equivalent de tres patates. Una persona «només [està formada per] un estómac i certs instints», ha escrit algú. Aquesta època, gravada encara a la memòria dels holandesos, va rebre el nom d’«hivern de la fam», o Hongerwinter.
La fam va persistir fins al 1945. Desenes de milers d’homes, dones i infants es van morir de desnutrició. Entre els milions de persones que van sobreviure, els canvis en l’alimentació van ser tan radicals i tan bruscos que van donar lloc a un experiment natural esgarrifós. Quan la població va sortir dels rigors hivernals, els investigadors van poder estudiar l’efecte d’aquella passa de fam sobre una cohort determinada de persones. S’hi van trobar algunes característiques previsibles, com la desnutrició i retards en el creixement. Els infants que van sobreviure al Hongerwinter també podien presentar problemes de salut crònics associats amb la desnutrició, com depressió, ansietat, malalties del cor, piorrea, osteoporosi i diabetis. (Audrey Hepburn, una actriu coneguda entre altres coses pel seu físic esprimatxat, va ser un d’aquests infants supervivents, i tota la vida va patir d’un gran nombre d’afeccions cròniques).
A la dècada del 1980, però, va sorgir un altre patró més intrigant: quan els fills d’algunes dones que estaven embarassades durant la crisi de fam van ser grans, van presentar uns índexs més elevats d’obesitat i de malalties del cor. Aquest descobriment, d’altra banda, s’hauria pogut preveure. Se sap que patir de desnutrició in utero causa alteracions en la fisiologia fetal. El fetus, mancat de nutrients, modifica el seu metabolisme per tal d’acumular quantitats més elevades de greixos i prevenir-se així de la pèrdua de calories; i això, paradoxalment, causa obesitat tardana i desordre metabòlic. El resultat més curiós de l’estudi del Hongerwinter, però, va trigar encara una generació més a descobrir-se. A la dècada del 1990, en què es va fer un estudi dels néts dels homes i les dones víctimes d’aquella crisi de fam, es va veure que també tenien índexs més alts d’obesitat i de malalties del cor (alguns d’aquests problemes de salut encara s’estan analitzant). La fase aguda d’inanició devia haver alterat de tal manera els gens en aquells qui l’havien patit directament que el missatge s’havia transmès als seus néts. En el genoma d’aquells homes i dones famolencs devia haver quedat imprès algun factor hereditari —o uns quants— que havia afectat almenys dues generacions. El Hongerwinter holandès no només havia quedat gravat en la memòria del país, sinó que també n’havia impregnat la memòria genètica.[070]
* * *
¿Què era la «memòria genètica», però? ¿Com es codificava la memòria dels gens més enllà dels gens mateixos? Waddington no va arribar a conèixer l’estudi sobre el Hongerwinter —s’havia mort el 1975 sense haver rebut pràcticament gens de reconeixement—, però els genetistes van saber veure la relació existent entre la hipòtesi de Waddington i les malalties multigeneracionals de la cohort holandesa. També en aquest cas semblava clar que hi havia una «memòria genètica»: els fills i els néts de persones que havien passat fam tendien a patir malalties metabòliques, com si al genoma hi tinguessin alguna mena de record de les penalitats metabòliques dels avis. També en aquest cas, el factor responsable del «record» no podia ser una alteració de la seqüència gènica, perquè era impossible que els centenars de milers d’homes i dones d’aquella cohort holandesa haguessin sofert tots una mutació en l’espai de tres generacions. I, també en aquest cas, una interacció «dels gens amb l’ambient» havia modificat un fenotip (la propensió a patir una malaltia). Alguna cosa devia haver quedat impresa en el genoma a causa de la fam —alguna mena de senyal permanent i hereditari— que s’estava transmetent d’una generació a la següent.
Si un plus d’informació així era susceptible de ser incorporat a un genoma, podia tenir conseqüències imprevisibles. En primer lloc, posava en qüestió un dels principis fonamentals de l’evolució darwiniana clàssica. Un element conceptualment cabdal de la teoria darwiniana és que els gens no recorden —no són capaços de recordar— cap experiència de l’organisme de forma heretable i permanent. Si un antílop estira el coll per arribar a un arbre alt, els seus gens no consignen aquest esforç, i les cries que tenen no són girafes (recordem que la transmissió directa d’una adaptació a un tret heretable era el fonament de l’errònia teoria de Lamarck de l’evolució a través de l’adaptació). En realitat les girafes apareixen arran de la variació espontània i la selecció natural: en un animal primigeni brostejador apareix un mutant de coll alt, i, durant una època de fam, aquest mutant sobreviu i és naturalment seleccionat. August Weismann havia contrastat formalment la hipòtesi segons la qual una influència ambiental podia alterar de forma permanent els gens tallant la cua de cinc generacions de ratolins, i els ratolins de la sisena generació havien nascut amb la cua absolutament intacta. L’evolució pot conformar organismes perfectament adaptats, però no de forma intencionada; no és només un «rellotger cec», com Richard Dawkins va definir-la amb encert, sinó també desmemoriat. Només la guien la supervivència i la selecció; com a record només té la mutació.
Tot i així, els néts del Hongerwinter semblaven haver adquirit el record de la fam que els seus avis havien passat, i no per mitjà de les mutacions ni la selecció, sinó d’un missatge ambiental que semblava que s’hagués tornat hereditari. Un «record» genètic d’aquest tipus es comportaria evolutivament com un «forat de cuc» (wormhole o pont d’Einstein-Rosen). Un avantpassat de la girafa seria capaç d’originar una girafa no pas avançant penosament per la feixuga lògica malthusiana de mutació, supervivència i selecció, sinó pel simple fet d’estirar el coll, cosa que deixaria consignat i gravat un record de l’estrebada en el genoma. Un ratolí amb la cua seccionada podria tenir ratolins amb la cua més curta perquè transmetria aquesta informació als gens. Els infants criats en ambients plens d’estímuls engendrarien infants més estimulats. Aquesta hipòtesi era una reformulació de la teoria darwiniana de les gèmmules: l’experiència o la història concretes d’un organisme quedaven gravades directament al genoma. Un sistema així es comportaria com una drecera entre l’adaptació i l’evolució de l’organisme. Tornaria la vista al rellotger.
Per a Waddington en concret, aquesta explicació tenia un interès afegit de caràcter personal. Era un partidari veterà i fervent del marxisme, i va pensar que el descobriment d’aquests tipus d’elements «fixadors de records» al genoma podien ser cabdals no només per comprendre el desenvolupament embriològic humà sinó també per al seu projecte polític particular. Si les cèl·lules podien ser adoctrinades o desadoctrinades per mitjà de la manipulació dels seus records genòmics, potser les persones també es podrien adoctrinar (recordem la temptativa de Lysenko d’aconseguir això mateix amb varietats de blat, i els propòsits d’Stalin de suprimir les ideologies de les persones dissidents). Un procés d’aquesta mena podia suprimir la identitat cel·lular i permetre que les cèl·lules ascendissin pel paisatge de Waddington, amb la qual cosa es passaria de nou de cèl·lula adulta a cèl·lula embrionària i s’invertiria el temps biològic. Fins i tot podria suprimir el caràcter fix de la memòria humana i de la identitat; és a dir, de la lliure elecció.
* * *
Fins a finals de la dècada del 1950 l’epigenètica va ser més una fantasia que no pas una realitat; ningú havia vist encara cap cèl·lula incorporant la seva història o la seva identitat al genoma. L’any 1961, dos experiments realitzats amb una diferència de menys de mig any, i a menys de trenta quilòmetres de distància l’un de l’altre, van transformar el coneixement dels gens i van donar credibilitat a la teoria de Waddington.
L’estiu del 1958, John Gurdon, un estudiant de doctorat de la universitat d’Oxford, va començar a estudiar el desenvolupament de les granotes. Gurdon no havia sigut fins aleshores un estudiant gaire prometedor —una vegada, en un examen de ciències, d’una classe de dos-cents cinquanta alumnes havia quedat l’últim—; però, com ell mateix va apuntar, tenia un «do per fer les coses a escala reduïda». El seu experiment més important no hauria pogut ser d’una escala més petita. A principis dels anys cinquanta, dos científics de Filadèlfia havien buidat de gens un òvul de granota sense fecundar per mitjà de la succió del nucli, i al material cel·lular que n’havia quedat, havien injectar el genoma d’una cèl·lula de granota d’un altre tipus. Era com buidar un niu i posar-hi un ocell que no corresponia per veure si s’hi desenvolupava normalment. ¿Disposava el niu (és a dir, l’òvul desposseït de tots els gens) de tots els elements necessaris per formar un embrió a partir del genoma d’una cèl·lula diferent? Va resultar que sí. D’un dels òvuls on havien injectat el genoma d’una cèl·lula diferent de granota, els investigadors de Filadèlfia van obtenir un capgròs. Era com una forma extrema de parasitisme: l’òvul esdevenia un simple hoste, o recipient, del genoma d’una cèl·lula normal, i feia possible que el genoma es transformés en un animal adult absolutament normal. Els dos investigadors van batejar el seu mètode amb el nom de transferència nuclear, però es tractava d’un procés molt poc eficaç i van acabar abandonant l’estudi.
Gurdon, fascinat per aquells resultats insòlits, va fer anar l’experiment més enllà. Els investigadors de Filadèlfia havien injectat nuclis d’embrions joves en els òvuls enucleats, però el 1961 Gurdon va començar a provar si la injecció del genoma d’una cèl·lula de l’intestí d’una granota adulta també engendrava un capgròs. Les dificultats tècniques eren enormes. En primer lloc, Gurdon va aprendre a utilitzar un feix molt prim de rajos ultraviolats per encertar el nucli d’un òvul de granota sense fecundar deixant el citoplasma intacte. A continuació, amb una agulla molt fina va fer una punció a la membrana de l’òvul sense pràcticament agitar-ne gens la superfície, com un bon saltador quan es capbussa a l’aigua, i hi va introduir el nucli d’una cèl·lula de granota adulta suspès en un volum ínfim de líquid.
La transferència d’un nucli de cèl·lula de granota adulta (és a dir, tots els seus gens) en un òvul buit va tenir èxit. En van néixer capgrossos vius, cada un dels quals contenia una còpia exacta del genoma de la granota adulta. Transferint els nuclis d’un conjunt de cèl·lules adultes procedents de la mateixa granota a un conjunt d’òvuls enucleats, Gurdon podia obtenir uns capgrossos que eren clons exactes entre ells, i alhora clons de la granota donant. Aquest procés es podia repetir ad infinitum, i anar obtenint clons de clons de clons, tots ells amb el mateix genotip exactament; és a dir reproduccions sense necessitat de reproducció.
L’experiment de Gurdon va despertar la imaginació dels biòlegs, entre altres coses perquè semblava una fantasia de ciència-ficció convertida en realitat. En un experiment va obtenir divuit clons a partir de cèl·lules intestinals d’una sola granota. Situats en divuit cambres idèntiques, eren com divuit dobles que visquessin en divuit universos paral·lels. El principi científic que es posava en qüestió també movia a reflexió: el genoma d’una cèl·lula adulta que havia arribat a la maduresa havia sigut submergit en el líquid d’un òvul i n’havia sortit absolutament rejovenit en forma d’embrió. L’òvul, per tant, tenia tot el que calia —tots els elements reguladors necessaris— per guiar un genoma enrere en el temps fins al primer estadi de desenvolupament embrionari. Amb el pas del temps, l’aplicació del mètode de Gurdon es va ampliar a altres espècies animals, fins a arribar a la cèlebre clonació de l’ovella Dolly, en aquells moments l’únic mamífer reproduït sense reproducció. (El biòleg John Maynard Smith va comentar que l’únic altre «cas conegut de mamífer engendrat sense sexualitat no era del tot convincent», referint-se a Jesucrist). L’any 2012, Gurdon va rebre el premi Nobel pel descobriment de la transferència nuclear.[071]
A desgrat de tots els aspectes notables de l’experiment de Gurdon, però, el més revelador va ser la seva falta d’èxit. Les cèl·lules intestinals de granotes adultes podien arribar a engendrar capgrossos, certament, però només gràcies a la meticulositat tècnica de Gurdon; l’índex d’èxit de conversió de cèl·lules adultes en capgrossos era molt baix. Això havia de tenir una explicació més enllà de la genètica clàssica. La seqüència d’ADN del genoma d’una granota adulta, al capdavall, és idèntica a la seqüència d’ADN d’un embrió de granota o d’un capgròs. El principi fonamental de la genètica, si més no, és que totes les cèl·lules contenen el mateix genoma, i que el que regula el desenvolupament de l’embrió fins a convertir-se en animal adult és la forma en què aquests gens s’expressen a les diferents cèl·lules, activats o reprimits d’acord amb unes pautes.
Si els gens són gens i res més, però, ¿per què al genoma d’una cèl·lula adulta li costava tant retrocedir fins a l’estadi d’embrió? ¿I per què, com van descobrir altres científics, els nuclis dels animals més joves eren més susceptibles a aquest rejoveniment que no els d’animals més vells? Igual que en l’estudi del Hongerwinter, també en aquest cas devia haver-hi alguna cosa que s’anava imprimint de mica en mica en el genoma de la cèl·lula adulta —alguna marca acumulativa i indeleble— que dificultava a aquell genoma de tornar enrere en el procés de desenvolupament. Aquesta marca no estava continguda pròpiament en la seqüència dels gens, sinó gravada a sobre; havia de ser epigenètica. Gurdon va tornar a centrar-se en la pregunta de Waddington: ¿podria ser que totes les cèl·lules tinguessin al genoma una empremta de la seva història i la seva identitat, una mena de memòria cel·lular?
* * *
Gurdon s’havia representat mentalment una marca epigenètica en sentit abstracte, però físicament no havia arribat a veure aquesta empremta al genoma de la granota. L’any 1961, Mary Lyon, antiga alumna de Waddington, va trobar un exemple visible d’un canvi epigenètic en una cèl·lula animal. Lyon, filla d’un funcionari i una mestra, va començar els estudis de doctorat a Cambridge amb Ron Fisher, cèlebre pel seu mal geni, però no va trigar gaire a fugir cap a Edimburg per acabar el doctorat, i posteriorment va anar a treballar en un laboratori de la plàcida població anglesa de Harwell, a una trentena de quilòmetres d’Oxford, on va crear el seu propi grup de recerca.
A Harwell, Lyon va estudiar la biologia dels cromosomes fent servir tincions químiques per distingir-los. Va tenir la sorpresa de descobrir que tots els cromosomes homòlegs tenyits amb tincions cromosòmiques es veien iguals tots dos a excepció de la parella de cromosomes X en els individus de sexe femení. En els ratolins femella, un dels dos cromosomes X apareixia sempre arronsat i tenyit de color més fosc. Els gens del cromosoma més dens i fosc no presentaven cap alteració, i la seqüència d’ADN era la mateixa en un cromosoma i l’altre. El que havia canviat entre ells era la seva activitat: els gens del cromosoma arronsat no sintetitzaven ARN, cosa que feia que tot el cromosoma fos «silenciós». Era com si un dels dos cromosomes hagués quedat fora de servei, apagat. Lyon va descobrir que l’elecció del cromosoma X inactivat havia sigut aleatòria; en una cèl·lula podia ser el cromosoma X masculí, mentre que a la cèl·lula del costat podia estar inactivat el cromosoma X femení. Es tractava d’un patró universal en totes les cèl·lules que tenien dos cromosomes X; és a dir, en totes les cèl·lules dels individus de sexe femení.
¿Quina funció compleix aquesta inactivació del cromosoma X? Com que els individus de sexe femení tenen dos cromosomes X, mentre que els de sexe masculí en tenen només un, les cèl·lules femenines inactiven un cromosoma X per compensar la «dosi» de gens dels dos cromosomes X. Aquesta inactivació aleatòria del cromosoma X té una conseqüència biològica important, que és que el cos dels individus de sexe femení és un mosaic de dos tipus de cèl·lules. Per regla general, aquest silenciament aleatori d’un cromosoma X és imperceptible, si no és que resulta que els cromosomes X (els paterns, posem per cas) contenen alguna variant gènica que produeix algun tret visible. En aquest cas, una cèl·lula pot expressar aquesta variant, mentre que la cèl·lula veïna pot estar mancada d’aquesta funció, cosa que crea un efecte de mosaic. En els gats, per exemple, al cromosoma X hi ha un gen del color del pèl. La inactivació aleatòria del comosoma X fa que una cèl·lula tingui un pigment de color i que la seva cèl·lula veïna tingui un color diferent. És l’epigenètica, no pas la genètica, el que aclareix l’enigma de les gates carei (tricolors). (Per exemple, si els éssers humans tinguéssim un únic gen del color de la pell localitzat al cromosoma X, la filla d’una parella de pell fosca i pell clara naixeria amb clapes de pell clara i clapes de pell fosca).
¿Com pot «silenciar» una cèl·lula tot un cromosoma? Aquest procés representa no només l’activació o la inactivació d’un o dos gens a partir d’algun estímul ambiental, sinó que un cromosoma sencer —amb tots els gens inclosos— queda inactivat durant tot el temps de vida de la cèl·lula. La hipòtesi més lògica, postulada a la dècada del 1970, era que les cèl·lules haguessin enganxat alguna mena de marca química permanent —un «mata-segells» molecular— a l’ADN d’aquell cromosoma. Com que els gens pròpiament dits quedaven intactes, aquesta empremta havia d’estar per sobre dels gens; és a dir, es tractava d’una marca epigenètica.
A finals de la dècada del 1970, uns científics que investigaven la silenciació gènica van descobrir que la unió d’una petita molècula —un grup metil— a algunes parts d’ADN presentava una correlació amb la inactivació d’un gen. Temps més tard es va saber que un dels inductors principals d’aquest procés és la molècula d’ARN produïda pel gen XIST.Aquesta molècula d’ARN «revesteix» parts del cromosoma i es creu que això és cabdal per al silenciament del cromosoma. Aquestes etiquetes de metil guarnien els filaments d’ADN com talismans en un collaret, i es va descobrir que constituïen senyals de silenciament d’alguns gens.
* * *
Les etiquetes de metil no eren els únics talismans que penjaven del collaret d’ADN. El 1996, mentre investigava a la universitat Rockefeller de Nova York, el bioquímic David Allis va trobar un altre mecanisme per deixar marques permanents en els gens. En comptes d’imprimir les marques en els gens directament, aquest altre mecanisme deixava les marques en unes proteïnes anomenades histones, que fan el paper de molècules empaquetadores de gens.[072]
Les histones formen una mena de petits nuclis (octàmers) que la cadena d’ADN embolcalla cargolant-s’hi com el fil en un rodet, cosa que crea una estructura d’empaquetament en què els nuclis d’histones embolcallats d’ADN serien com els grans d’un rosari. Quan hi ha algun canvi en aquest procés d’empaquetament, l’activitat d’un gen pot canviar, tal com passa quan les propietats d’un material s’alteren segons la forma en què està embalat (un fil de seda en troca, per exemple, té propietats molt diferents de quan està descargolat i estirat). Podria imprimir-se una «memòria molecular» en un gen, però de manera indirecta, deixant el senyal en una proteïna. (Dintre del camp de l’epigenètica hi ha un gran debat sobre si la modificació de les histones té algun efecte rellevant sobre l’activitat dels gens o si aquests canvis en les histones són simples «testimonis» o efectes secundaris de l’activitat d’un gen). Encara s’està investigant sobre l’estabilitat i l’heretabilitat d’aquestes empremtes en les histones, i sobre el mecanisme que vetlla perquè aquestes empremtes apareguin en els gens adequats i el moment adequat, però sembla que els organismes simples com els llevats i els cucs conserven la capacitat de transmetre-les al llarg d’unes quantes generacions.
* * *
El silenciament i l’activació dels gens per mitjà dels reguladors proteics coneguts amb el nom de factors de transcripció —els «directors d’orquestra» de la simfonia de gens de les cèl·lules— són coneguts des de la dècada del 1950. Aquests directors, però, poden recórrer a altres proteïnes —que en podríem dir ajudants— perquè deixin marques químiques permanents en els gens. Fins i tot poden arribar a vetllar perquè les etiquetes es conservin al genoma.[073] Això permet que les etiquetes es puguin afegir, esborrar, ampliar, reduir i activar o desactivar en funció de senyals cel·lulars o ambientals.[074]
Aquestes marques funcionen com notes escrites damunt d’una frase, o com postil·les en un llibre —marques amb llapis, paraules subratllades, guixots, lletres ratllades, comentaris al peu, notes finals—, que modifiquen el context del genoma sense canviar-ne les paraules. Totes les cèl·lules d’un organisme hereten el mateix llibre; però ratllant unes frases i afegint-n’hi d’altres, «silenciant» i «activant» paraules concretes, destacant segons quines frases, cada cèl·lula podria arribar a escriure una novel·la única a partir d’un guió comú. Podríem veure els gens del genoma humà, amb les seves marques químiques incorporades, de la manera següent:
… . Aquesta … … és … . l’ … … … , , , … … … .estruc … . tura , … … del … .Nostre … … … Gen … . oma … .
Igual que l’altra vegada, les paraules de la frase corresponen als gens. Els punts suspensius i els signes de puntuació indiquen els introns, les regions intergèniques i les sequències reguladores. Les lletres en negreta i en majúscules i les paraules subratllades són marques epigenètiques afegides al genoma que hi aporten un matís de significat particular.
És precisament per aquest motiu que Gurdon, tot i la seva meticulositat tècnica, només comptades vegades havia aconseguit induir una cèl·lula intestinal adulta de granota a tornar enrere en el seu procés de desenvolupament fins a arribar a l’estadi embrionari, i, posteriorment, a convertir-se en una granota adulta: el genoma de la cèl·lula intestinal estava ja massa marcada amb notes «epigenètiques» perquè s’esborressin fins a transformar-se en un genoma d’embrió. Igual que els records de les persones, que persisteixen per més que se’ls vulgui alterar, aquests gargots químics sobreescrits al genoma costen molt de modificar. Aquests senyals estan dissenyats per ser persistents, de tal manera que una cèl·lula pugui consolidar la seva identitat. Únicament les cèl·lules embrionàries tenen genomes susceptibles d’adquirir molts tipus d’identitat diferents i, així, engendrar tots els tipus cel·lulars del cos. Quan les cèl·lules embrionàries ja han adquirit una identitat determinada —és a dir, quan ja s’han convertit en cèl·lules intestinals, sanguínies o nervioses, per exemple— rara vegada hi ha marxa enrere. (Per això a Gurdon li va costar tant obtenir un capgròs a partir d’una cèl·lula intestinal de granota). Una cèl·lula embrionària és capaç d’escriure una infinitat de novel·les a partir del mateix guió; però un relat juvenil, un cop escrit, difícilment es pot reconvertir en una novel·la victoriana.
* * *
La interacció entre els gens reguladors i l’epigenètica explica en part l’enigma de la individualitat de les cèl·lules, i fins i pot podria arribar a explicar l’enigma encara més impenetrable de la individualitat de l’individu. Més amunt hem formulat la pregunta de per què els bessons són diferents. Doncs ho són perquè uns fets singulars els queden gravats al cos en forma de marques singulars. Ara bé, ¿«gravats» de quina manera? No pas en la seqüència gènica pròpiament dita; perquè, si seqüenciem els genomes d’una parella de bessons univitel·lins cada deu anys durant cinquanta anys, cada vegada obtindrem la mateixa seqüència. En canvi, si seqüenciem els epigenomes d’una parella de bessons al llarg d’unes quantes dècades, hi trobarem diferències substancials. El patró de grups metil que hi ha enganxats als genomes de les cèl·lules sanguínies o de les neurones, que és pràcticament idèntic a tots dos bessons al principi de l’experiment, comença a divergir de mica en mica durant els primers deu anys, i al cap de cinquanta anys és substancialment diferent.[075]
Certs esdeveniments fortuïts —ferides, infeccions, enamoraments; el trinat inoblidable d’aquell nocturn concret; l’olor d’aquella magdalena concreta a París— afecten un dels bessons però no l’altre. Les proteïnes reguladores activen o reprimeixen gens com a reacció a aquests esdeveniments, i les marques epigenètiques es van acumulant en els gens de mica en mica.[076] Encara no s’ha establert de quina manera aquestes marques epigenètiques influeixen a la pràctica en l’activitat dels gens; però hi ha experiments que indiquen que aquestes marques, en conjunció amb els factors de transcripció, poden ajudar a organitzar l’activitat dels gens.
Al seu memorable relat «Funes el memorioso», l’escriptor argentí Jorge Luis Borges descriu un jove que, quan després d’un accident recobra els sentits, descobreix que ha adquirit una memòria «infal·lible». Funes recorda tots els detalls de tots els moments de la seva vida, tots els objectes, totes les coneixences que ha fet; «les formes dels núvols […] [i] les vetes d’un llibre». Aquesta capacitat extraordinària, però, no converteix Funes en algú més poderós, sinó que el paralitza. L’inunda un devessall de records que no pot reprimir; els records l’atueixen, com la remor constant d’una multitud que no pots fer callar. El narrador troba Funes estirat en un catre, a les fosques, incapaç de contenir aquella allau terrible d’informació i obligat a mantenir-se apartat del món.
Una cèl·lula sense capacitat de reprimir selectivament parts del seu genoma esdevé un Funes memoriós (o, com al relat, un Funes impossibilitat). El genoma conté la memòria per crear qualsevol cèl·lula de qualsevol teixit de l’organisme; és una memòria tan rica i diversa que una cèl·lula mancada d’un sistema de repressió i reactivació selectives queda atuïda de tanta riquesa i diversitat. Tal com li passa a Funes, paradoxalment, la capacitat d’utilitzar a la pràctica qualsevol memòria depèn de la capacitat de reprimir memòria. Potser el sistema epigenètic és necessari perquè el genoma pugui complir la seva funció. Encara hi ha moltes coses per descobrir-ne. Pel que sembla, en cèl·lules diferents el genoma pot ser modificat diferencialment per marques químiques diverses com a reacció a estímuls diversos (incloent-hi les condicions ambientals). En tot cas, la possibilitat que aquestes marques puguin contribuir a l’activitat dels gens, la forma com pot ser que ho facin, i quina funció deuen complir, continuen sent motiu de debats encesos i aferrissats entre els genetistes.
* * *
La demostració segurament més colpidora de la capacitat de les proteïnes mestres reguladores per dirigir l’orquestra de les marques epigenètiques amb la finalitat de reinicialitzar la memòria cel·lular és un experiment realitzat el 2006 pel biòleg japonès especialista en cèl·lules mare Shinya Yamanaka. Igual que Gurdon, Yamanaka estava intrigat per la possibilitat que les marques químiques unides als gens d’una cèl·lula poguessin tenir la funció de definir la identitat cel·lular. ¿Què passaria si aconseguia esborrar aquestes marques? ¿Podria passar que la cèl·lula adulta tornés a un estat anterior i es reconvertís en una cèl·lula embrionària, en una mena d’inversió del temps en què la història quedés abolida i es retornés a l’estat més primerenc?
També igual que Gurdon, Yamanaka va començar el seu projecte de fer perdre la identitat a una cèl·lula amb una cèl·lula normal d’un ratolí adult, en aquest cas del teixit epitelial. L’experiment de Gurdon havia demostrat que certs factors presents a l’òvul —proteïnes i ARN— podien esborrar les marques del genoma d’una cèl·lula adulta i per tant invertir l’especificació de la cèl·lula, de tal manera que es podia obtenir un capgròs a partir d’una cèl·lula de granota. Yamanaka es va proposar d’identificar i aïllar aquests factors de l’òvul, per després utilitzar-los com a «goma d’esborrar» de l’especificació cel·lular. Després de dècades de recerca, Yamanaka va aconseguir identificar aquests factors misteriosos com unes proteïnes codificades per quatre gens concrets, i posteriorment va introduir aquests quatre gens en cèl·lules epitelials de ratolí adult.
Yamanaka va tenir una sorpresa que es va convertir en admiració entre els científics de tot el món: la introducció d’aquells quatre gens en cèl·lules epitelials madures va provocar que una petita part de les cèl·lules en transformés en un tipus cel·lular semblant a cèl·lules mare embrionàries. Aquestes cèl·lules mare eren capaces de formar cèl·lules epitelials, per descomptat, però també musculars, òssies, intestinals i nervioses; eren capaces de donar origen a qualsevol tipus cel·lular existent a l’organisme. Yamanaka i els seus col·legues van analitzar la progressió (o més aviat regressió) de les cèl·lules epitelials fins a l’estat de cèl·lula pseudoembrionària, i van posar al descobert tot un seguit de fets. S’activaven o es reprimien circuits de gens. Es reinicialitzava el metabolisme de la cèl·lula. A continuació s’esborraven i es reescrivien les marques epigenètiques. La cèl·lula canviava de forma i de mida. Li desapareixien les arrugues, recuperava l’elasticitat de les articulacions encarcarades, recobrava la joventut i era capaç d’ascendir pel paisatge en pendent de Waddington. Yamanaka havia eliminat la memòria d’una cèl·lula, havia invertit el rellotge biològic.
Però es tractava d’un regal enverinat. Un dels quatre gens utilitzats per Yamanaka per invertir l’especificitat cel·lular es diu c-myc. Myc, el factor del rejoveniment, no és un gen normal i corrent, sinó que és un dels reguladors més potents del creixement i el metabolisme cel·lulars que es coneixen en biologia. Si se l’activa de forma anòmala fa retrocedir una cèl·lula adulta fins a un estat pseudoembrionari, tal com s’ha explicat, i per tant fa possible l’experiment d’inversió de l’especificitat cel·lular de Yamanaka (una funció que exigeix la col·laboració dels altres tres gens descoberts per Yamanaka). Ara bé, myc també és un dels proto-oncogens cel·lulars més potents que es coneixen en biologia, que també s’activa en les leucèmies i els limfomes i en els càncers de pàncrees, d’estómac i d’úter. Com si es tractés d’una faula moral antiga, sembla que la recerca de l’eterna joventut té un cost col·lateral terrible. Els mateixos gens que fan possible que una cèl·lula es desfaci de la mortalitat i l’envelliment també poden encaminar el seu destí cap a la immortalitat mortífera, el creixement imparable i la incapacitat d’envellir que són els trets distintius del càncer.
* * *
Ara ja podem mirar de comprendre el Hongerwinter holandès, i els seus efectes al llarg de generacions, en uns termes mecanicistes que involucren tant els gens com les proteïnes reguladores que interactuen amb el genoma. La inanició aguda que van patir tant homes com dones durant aquells mesos atroços de l’any 1945 va alterar sens dubte l’expressió dels gens relacionats amb el metabolisme i l’emmagatzematge. Els primers canvis van ser transitoris, segurament poca cosa més que una activació i inactivació dels gens que reaccionen en funció del nivell de nutrients disponibles.
Quan la inanició es va fer crònica, però, el programa metabòlic es va «congelar» i reinicialitzar —l’estat transitori es va consolidar com a permanent—, i al genoma hi van quedar gravats canvis més persistents. Les hormones es van difondre pels òrgans per advertir de la possible prolongació de la falta d’aliment i d’una reconversió més àmplia de l’expressió gènica. Les proteïnes van rebre aquests missatges a l’interior de les cèl·lules. Els gens es van anar inactivant un per un, i a l’ADN es van posar les empremtes que refermaven la inactivació. De la mateixa manera que la gent tapa totes les obertures de les cases quan s’acosta una tempesta, es van tancar amb pany i forrellat programes gènics complets. Es van afegir marques de metilació als gens. Segurament les histones es van modificar químicament per deixar l’empremta de la inanició.
Cèl·lula a cèl·lula, òrgan a òrgan, el cos va ser reprogramat per sobreviure. Finalment, fins i tot les cèl·lules germinals —precursores de l’òvul i l’espermatozou— van quedar marcades. (No sabem com ni per què, però el fet és que l’òvul i l’espermatozou contenen el record de la reacció a la inanició; potser en el genoma hi ha antics mecanismes que registren en les cèl·lules mare els episodis d’inanició o de penúria).[077] Posteriorment, els embrions formats a partir d’aquests òvuls i espermatozous —els futurs fills i néts d’aquelles dones i aquells homes— devien contenir aquestes marques, perquè van acabar presentant alteracions metabòliques que s’han mantingut gravades al seu genoma dècades després del Hongerwinter. La memòria històrica havia esdevingut memòria cel·lular.
* * *
Un advertiment: l’epigenètica també corre el perill d’esdevenir una idea perillosa. Teòricament, les modificacions epigenètiques dels gens poden superposar informació històrica i ambiental en les cèl·lules i els genomes, però aquesta capacitat és hipotètica, limitada, singular i imprevisible: un progenitor que ha patit d’inanició engendra fills amb obesitat i hipernutrició, però un pare que ha tingut la tuberculosi no té fills amb una resposta alterada a la tuberculosi. La majoria de la «memòria» epigenètica és producte d’antics mecanismes evolutius, i no s’han de confondre amb el nostre desig de deixar una bona herència als fills.
Tal com va passar amb la genètica a principis del segle XX, actualment l’epigenètica es fa servir per justificar les pseudociències i per imposar definicions ràncies de normalitat. Les dietes, els rajos X, els records i les teràpies que pretesament alteren l’herència recorden inquietantment el projecte de Lysenko de «reeducar» el blat per mitjà d’una teràpia de xoc. S’aconsella a les mares que procurin no passar ansietat durant l’embaràs per tal de no deixar cap estigma en els fills, ni en els fills dels seus fills, amb forma de mitocondri traumatitzat. Lamarck retorna com un nou Mendel.
Aquestes idees simplistes sobre l’epigenètica haurien de convidar-nos a l’escepticisme. És cert que la informació procedent del medi es pot gravar al genoma; però la majoria d’aquestes empremtes queden registrades com a «memòria genètica» a les cèl·lules i als genomes d’organismes individuals, i no es transmeten a les generacions següents. Un home que perd una cama en un accident té l’empremta de l’accident a les cèl·lules, les ferides i les cicatrius, però no té fills camacurts. I tampoc la vida expatriada de la meva família sembla haver-nos carregat, ni a mi ni als meus fills, amb cap sentiment punyent d’enyorança.
A desgrat de l’afirmació de Menelau, la sang dels nostres pares sí que s’ha extingit en nosaltres; i igualment, per sort, les seves febleses i les seves faltes. És un estat de coses que no hauríem de lamentar, sinó de celebrar. Els genomes i els epigenomes existeixen per registrar i transmetre semblança, herència, memòria i història a les cèl·lules i a les successives generacions. Les mutacions, la reordenació de gens i la supressió de la memòria epigenètica contraresten aquestes forces i fan possibles la diferència, la variació, l’aberració, el geni i la reinvenció, i per tant l’esplendorosa possibilitat de començar de nou una generació rere l’altra.
* * *
És molt versemblant que la interacció de gens i epigens coordini l’embriogènesi humana. Retornem un moment al problema de Morgan, que és la formació d’un organisme pluricel·lular a partir d’un embrió unicel·lular. Pocs segons després de la fecundació, a l’embrió comença a haver-hi moviment. Les proteïnes entren al nucli de la cèl·lula i comencen a activar i desactivar interruptors genètics. Una nau espacial adormida cobra vida. Els gens són activats i reprimits, i aquests mateixos gens, per la seva banda, codifiquen unes altres proteïnes que activen i reprimeixen uns altres gens. Una sola cèl·lula es divideix primer en dues cèl·lules, després en quatre, després en vuit. Es forma tota una capa de cèl·lules, que tot seguit es bufa com el cuiro d’una pilota quan la infles. Els gens que coordinen el metabolisme, la mobilitat, l’especificació cel·lular i la identitat es «disparen». La sala de màquines s’engega. Els llums fan pampallugues als corredors. L’intercomunicador es desperta entre espetecs.
Tot seguit entra en acció una nova tongada d’informació —induïda per proteïnes reguladores principals— que fa que l’expressió gènica quedi fixada a cada cèl·lula, cosa que permet que cada cèl·lula adquireixi i consolidi una identitat. S’afegeixen selectivament marques químiques a uns gens i se n’esborren d’altres, fet que regula l’expressió gènica d’aquella cèl·lula en concret. Es conjuguen i desconjuguen grups metil, i les histones es modifiquen per activar o reprimir gens.
L’embrió es desenvolupa pas a pas. Tenen lloc els primers estadis de segmentació, i les cèl·lules ocupen el seu lloc a les diferents parts de l’embrió. S’activen nous gens que executen subprogrames per formar les extremitats i els òrgans, i s’afegeixen més marques químiques específiques als genomes de cada tipus de cèl·lula. Les cèl·lules es multipliquen per formar òrgans i estructures: les extremitats anteriors, les extremitats posteriors, la musculatura, els ronyons, els ossos, els ulls. Algunes cèl·lules es moren per un procés de mort cel·lular programada. Els gens encarregats de preservar la funció, el metabolisme i el manteniment cel·lular s’activen. D’una cèl·lula sorgeix un organisme.
* * *
Que aquesta descripció no ens atabali. No tinguem la temptació de pensar «Quina fórmula més complicada, Déu meu!» i quedar tranquils tot seguit, convençuts que ningú serà capaç d’entendre, sabotejar ni manipular aquesta fórmula amb algun propòsit concret.
Quan els científics subestimen la complexitat, s’exposen als perills de les conseqüències imprevistes. Els exemples d’aquesta mala actitud són coneguts: animals foranis introduïts per controlar plagues que es converteixen en una plaga ells mateixos; l’augment de l’alçada de les xemeneies feta per reduir la contaminació urbana allibera gasos residuals més enlaire i fa augmentar la contaminació; l’estimulació de la síntesi de sang, que té l’objectiu de prevenir els atacs de cor, espesseix la sang i augmenta el risc de formació de coàguls sanguinis al cor.
Ara bé, quan els llecs sobreestimen la complexitat —«No pot ser que algú sigui capaç de desxifrar aquest codi»— cauen en la trampa de les conseqüències inesperades. A principis de la dècada del 1950, un tòpic corrent entre alguns biòlegs era que el codi genètic havia de ser tan dependent de l’entorn —tan determinat per una cèl·lula concreta d’un organisme concret, i tan tremendament complicat— que desxifrar-lo seria impossible. La realitat ho va desmentir completament: el codi està contingut en únic tipus de molècula, i es tracta d’un codi únic per a tot el món dels éssers vius. Coneixent el codi, podem alterar-lo a voluntat en els organismes en general, també en l’ésser humà. Una cosa semblant va passar a la dècada del 1960, en què molta gent posava en dubte que les tècniques de clonació genètica fossin capaces de transferir gens d’una espècie a una altra. Pel volts del 1980, introduir una proteïna de mamífer en una cèl·lula bacteriana, o una proteïna bacteriana en una cèl·lula de mamífer, no només era factible, sinó, com va dir Berg, «ridículament simple». Les espècies eren especioses. «La naturalitat» sovint no era «res més que façana».
És innegable que la gènesi d’un ésser humà a partir d’instruccions genètiques és complexa, però no té res que impedeixi ni limiti la manipulació o la modificació. Quan un sociòleg afirma que el que determina la forma, la funció i el comportament no són els gens tots sols, sinó les interaccions entre els gens i l’ambient, subestima la influència dels gens mestres reguladors, que actuen de forma autònoma i no condicionada per determinar estats fisiològics i anatòmics complexos. I quan un genetista diu que «la genètica no es pot fer servir per manipular estats i comportaments complexos perquè aquests estats acostumen a estar regulats per molts gens alhora», aquest genetista subestima la capacitat d’un únic gen, com per exemple un gen mestre regulador, de «reinicialitzar» estats d’existència complets. Si l’activació de quatre gens és capaç de convertir una cèl·lula epitelial en una cèl·lula mare pluripotent, si un fàrmac és capaç de revertir la identitat d’un cervell, i si una mutació d’un sol gen és capaç de canviar el sexe i la identitat de gènere, vol dir que tant el nostre genoma com la nostra persona són molt més mal·leables del que ens havíem imaginat.
* * *
La tecnologia, com he dit més amunt, és especialment eficaç quan fa possible un canvi o transició: del moviment lineal al moviment circular (la roda), o de l’espai real a l’espai virtual (internet). La ciència, en canvi, és especialment eficaç quan dilucida regles d’organització —lleis— que actuen com prismes a través dels quals es pot observar i ordenar el món. Els tecnòlegs, per mitjà d’aquelles transicions, tracten de deslliurar-nos de les limitacions de la nostra realitat present, mentre que els científics defineixen aquestes limitacions i tracen la frontera de l’horitzó de les coses possibles. Per això és que les innovacions tecnològiques més importants que hi ha hagut porten noms que donen testimoni del nostre domini sobre el món: l’enginyeria (derivat d’enginy, de la família de geni) o la computació (parent culte de comptar). Les lleis científiques més cabdals, en canvi, són batejades sovint amb termes que expressen els límits del coneixement humà: incertesa, relativitat, incompletesa, impossibilitat.
De totes les ciències, la biologia és la més anàrquica; en primer lloc, hi ha poques regles, i encara n’hi ha menys que siguin universals. És clar que els éssers vius obeeixen les regles fonamentals de la física i la química, però la vida acostuma a existir als marges i als intersticis d’aquestes lleis, i les força quasi fins al límit. L’univers busca equilibris; tendeix a dispersar l’energia, desbaratar l’organització i augmentar el caos. La vida, en canvi, està concebuda per combatre aquestes forces. Alentim les reaccions, concentrem la matèria i disposem les substàncies químiques en compartiments; mantenim l’ordre. «A vegades sembla que l’objectiu quixotesc que tenim en aquest món sigui refrenar l’entropia», ha escrit James Gleick. Vivim a les escletxes de les lleis naturals buscant extrapolacions, excepcions i excuses. Les lleis de la naturalesa no deixen d’assenyalar els límits del que és permès; però la vida, amb tota la seva estrafolària singularitat, prospera llegint el reglament entre línies. Ni tan sols l’elefant pot violar les lleis de la termodinàmica; però la trompa que té deu figurar entre els mitjans més peculiars de moure matèria per mitjà de l’energia.
* * *
Aquest és el flux circular de la informació biològica:
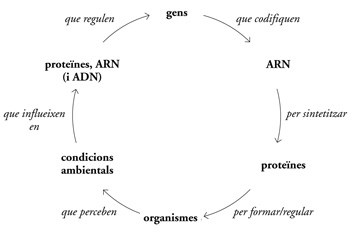
Segurament és una de les poques regles organitzatives que hi ha en biologia. És cert que la direccionalitat d’aquest flux d’informació té excepcions (els retrovirus poden «pedalar enrere» i obtenir ADN a partir d’ARN), i que en el món dels éssers vius encara hi ha mecanismes per descobrir que podrien modificar l’ordre o els elements del flux d’informació en els sistemes biològics (actualment se sap, per exemple, que l’ARN té la capacitat d’influir sobre la regulació dels gens), però el flux circular de la informació biològica queda aquí esbossat com a concepte.
Aquest flux d’informació és la cosa més pròxima a una llei biològica de què podem disposar. Quan dominem la tecnologia per manipular aquesta llei farem una de les transicions més importants de la nostra història. Aprendrem a llegir i escriure les nostres persones; aprendrem a llegir-nos i a escriure’ns.
* * *
Abans de fer aquest salt al genoma del futur, però, m’agradaria fer una ràpida digressió sobre el seu passat. No sabem d’on vénen els gens ni com van sorgir, i tampoc sabem per què precisament va ser seleccionat aquest mètode de transmissió d’informació i d’emmagatzematge de dades d’entre tots els altres possibles mètodes que hi ha en biologia; però podem provar de reconstruir en un tub d’assaig l’origen primordial dels gens. A Harvard, un bioquímic de parlar melós que es diu Jack Szostak s’ha passat més de vint anys intentant obtenir en un tub d’assaig un sistema genètic amb capacitat reproductiva, és a dir, reconstruint l’origen dels gens.
L’experiment de Szostak és posterior als estudis d’Stanley Miller, el químic visionari que havia intentat fer un «brou primordial» barrejant compostos químics que se sabia que eren presents a l’antiga atmosfera. A la dècada del 1950, en què treballava a la universitat de Chicago, Miller havia tancat hermèticament un baló de vidre i hi havia insuflat metà, diòxid de carboni, amoníac, oxigen i hidrogen per mitjà d’un conjunt de conductes. A aquesta mescla hi havia afegit vapor d’aigua calent i hi havia instal·lat un elèctrode per simular llampecs, i s’havia dedicat a escalfar i refredar alternativament el baló per reproduir les condicions inestables del món primigeni. En un matràs rodó hi havia ajuntat sofre i foc, cel i infern, aire i aigua.
Passades tres setmanes, del baló de Miller no havia sortit cap organisme; però en aquella barreja tosca de diòxid de carboni, metà, aigua, amoníac, oxigen, hidrogen, calor i electricitat, Miller hi havia trobat traces d’aminoàcids —les unitats estructurals de les proteïnes— i petites quantitats dels sucres més simples. En subsegüents variacions de l’experiment, Miller hi havia afegit argila, basalt i roca volcànica, i havia obtingut àcids grassos, els elements constituents dels lípids, i fins i tot les unitats bàsiques de l’ARN i l’ADN.
Szostak creu que els gens van sorgir d’aquest brou gràcies a la trobada fortuïta de dos insòlits companys de viatge. En primer lloc, els lípids que es van formar en el brou es van aglomerar entre ells i van formar micel·les, una mena de membranes esfèriques buides parentes de les bombolles de sabó que contenen líquid a dintre i s’assemblen a les membranes de les cèl·lules (Hi ha certs greixos que, barrejats a l’interior d’una solució aquosa, tendeixen a aglomerar-se espontàniament i a formar aquesta mena de bombolles). Amb els seus experiments de laboratori, Szostak ha demostrat que aquestes micel·les es poden comportar com protocèl·lules: si se’ls subministren més lípids, aquestes «cèl·lules» buides comencen a augmentar de mida. Es dilaten, es mouen i formen unes protuberàncies primes semblants a les ondulacions de les membranes cel·lulars. Al final acaben dividint-se, i de cada micel·la en surten dues.
En segon lloc, mentre aquestes micel·les s’anaven formant totes soles, van sorgir cadenes d’ARN a partir de la unió de nucleòtids (A, C, G, U o els seus predecessors químics). La major part d’aquestes cadenes d’ARN no tenien capacitat reproductiva; no podien fer còpies de si mateixes. Entre aquests milers de milions de molècules d’ARN incapaces de reproduir-se, però, casualment en va aparèixer una amb la capacitat exclusiva de fabricar una imatge de si mateixa; o, més ben dit, de fer una còpia per mitjà de la seva pròpia imatge especular. (Recordem que l’ARN i l’ADN tenen una estructura química intrínseca que fa possible la creació de molècules especulars). La molècula d’ARN, encara que pugui semblar mentida, posseïa la capacitat d’obtenir nucleòtids d’una barreja química i lligar-los entre ells per formar una còpia d’ARN nova. Era un compost químic amb la capacitat de duplicar-se.
El pas següent va ser un matrimoni de conveniència. En algun lloc del planeta —Szostak creu que devia ser a la riba d’un estany o d’un terreny pantanós— una molècula d’ARN amb capacitat reproductiva va coincidir amb una micel·la amb capacitat reproductiva. Com a concepte, va ser una bomba: les dues molècules es van conèixer, es van enamorar i van iniciar una llarga relació conjugal. L’ARN capaç de replicar-se va començar a viure a l’interior de la micel·la capaç de dividir-se. La micel·la es va recloure per protegir l’ARN, i dintre de la seguretat de la bombolla s’hi van poder produir determinades reaccions químiques. La molècula d’ARN, per la seva banda, va començar a codificar informació que resultava profitosa no només per reproduir-se ella sola, sinó conjuntament amb la micel·la. Amb el temps, la informació codificada al complex de l’ARN i la micel·la va fer possible crear més complexos de micel·la amb ARN.
«És relativament fàcil imaginar-se com devien evolucionar aquestes protocèl·lules amb ARN», ha escrit Szostak. «El metabolisme es devia anar ampliant de mica en mica, a mesura que […] [les protocèl·lules aprenien a] sintetitzar nutrients pel seu compte a partir de matèries primeres més simples i més abundants. A continuació, els organismes devien afegir la síntesi de proteïnes al seu utillatge químic». Els «protogens» d’ARN devien aprendre a induir els aminoàcids a formar cadenes, és a dir, a sintetitzar proteïnes, uns versàtils artefactes moleculars que dotaven de molta més eficàcia el metabolisme, la reproducció i la transmissió d’informació.
* * *
¿Quan i per què van aparèixer «gens» diferenciats (mòduls d’informació) en un filament d’ARN? Els gens ¿existien en la seva forma modular de bon començament o bé hi va haver alguna forma intermèdia o alternativa d’emmagatzematge d’informació? Una vegada més, es tracta de preguntes impossibles de respondre; però potser la teoria de la informació ens pot proporcionar alguna pista important. El problema que té la informació contínua, no modular, és que costa moltíssim treballar-hi. Tendeix a disgregar-se, a corrompre’s; tendeix a barrejar-se, adulterar-se i perdre’s. Si en tibes una punta es descabdella de l’altra. Si la informació té interferències internes, hi ha un risc molt més gran de distorsió; pensem per exemple en un disc de vinil que tingui una ratllada just al mig. La informació «digitalitzada», en canvi, és molt més fàcil de reparar i de recuperar. Tenim la possibilitat d’accedir a una paraula concreta d’un llibre i canviar-la sense haver de canviar la distribució de tota la biblioteca. Els gens haurien pogut aparèixer pel mateix motiu, com a mòduls diferenciats en un filament d’ARN que contenen informació destinada a codificar instruccions per portar a terme funcions diferenciades i concretes.
La naturalesa discontínua de la informació devia aportar un avantatge afegit: una mutació podia afectar un gen, però un i prou, i no els altres. Les mutacions podien actuar, per tant, sobre mòduls d’informació específics, sense pertorbar la funció de l’organisme com un tot, i així podien accelerar l’evolució. Aquest avantatge, però, venia acompanyat d’un perill: si hi havia més mutacions del compte, la informació podia deteriorar-se o perdre’s. El que convenia era tenir una còpia de seguretat, una imatge especular per protegir l’original o per restaurar-lo si es feia malbé. Segurament aquesta va ser la causa principal que va propiciar l’aparició d’un àcid nucleic de doble cadena. Les dades d’una de les cadenes quedaven perfectament reproduïdes en l’altra i es podien fer servir per restaurar qualsevol deteriorament; el yin protegia el yang. Així va ser com la vida es va inventar el seu disc dur particular.
Amb el temps, aquesta còpia nova —l’ADN— es va acabar convertint en l’original. L’ADN va ser una invenció del món de l’ARN, però no va trigar a superar l’ARN com a portador de gens, fins a convertir-se en el dipositari principal de la informació genètica en els sistemes vius.[078] En la història dels nostres genomes, doncs, encara hi ha gravat un altre mite de l’antiguitat: el del fill que es menja el pare, o Cronos destronat per Zeus.