Regulació, replicació, recombinació
Necessité absolue trouver origine de cet emmerdement [Cal descobrir com sigui l’origen d’aquest embolic]
JACQUES MONOD
De la mateixa manera que la formació d’un cristall gegant es pot induir disposant ordenadament uns quants àtoms determinats al seu nucli, el naixement d’un gran corpus científic es pot materialitzar a partir de la interrelació d’unes quantes idees fonamentals. Abans de Newton, unes quantes generacions de físics havien reflexionat sobre fenòmens com la força, l’acceleració, la massa i la velocitat. La genialitat de Newton va consistir a definir aquests conceptes de forma rigorosa i a relacionar-los entre ells per mitjà d’un teixit d’equacions que va fundar la ciència de la mecànica.
Seguint una lògica semblant, la interrelació d’unes quantes idees fonamentals va refundar la ciència de la genètica:
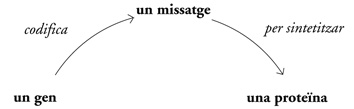
Tal com havia passat amb la mecànica newtoniana, amb el pas del temps el «dogma central» de la genètica es va depurar, modificar i reformular enormement. El seu efecte sobre la nova ciència, però, va ser profund, perquè va fixar un sistema de pensament. L’any 1909, Johannsen, en el moment d’encunyar el terme gen, l’havia declarat «lliure de tota hipòtesi». A principis de la dècada del 1960, però, el gen havia passat a ser molt més que una «hipòtesi». La genètica havia trobat una manera d’explicar la transmissió d’informació d’un organisme a un altre; i, dintre d’un organisme, el pas del codi xifrat a la forma. S’havia posat al descobert el «mecanisme» de l’herència.
Ara bé, ¿com podia ser que aquesta transmissió d’informació biològica assolís la complexitat que s’observava en els organismes vius? Prenguem com a exemple el cas de l’anèmia falciforme. Walter Noel havia heretat dues còpies anòmales del gen de l’hemoglobina B. Totes les cèl·lules del seu cos contenien les dues còpies anòmales (perquè totes les cèl·lules del cos posseeixen el mateix genoma). Les úniques cèl·lules que Noel tenia afectades pels gens alterats, però, eren els glòbuls vermells, no les neurones ni les cèl·lules dels ronyons, del fetge o de la musculatura. ¿Què permetia l’«acció» selectiva de l’hemoglobina en els glòbuls vermells? ¿Per què Noel no tenia hemoglobina a l’ull ni a la pell, per més que les cèl·lules de l’ull i de la pell, i de fet totes les cèl·lules del cos, tinguessin còpies idèntiques del mateix gen? Dit amb les paraules de Thomas Morgan: ¿per què «les propietats implícites en els gens es fan explícites en cèl·lules [diferents]»?
* * *
L’any 1940, un experiment realitzat amb un dels organismes més simples —Escherichia coli, un bacteri microscòpic amb forma de càpsula que viu als budells— va proporcionar la primera pista important sobre aquesta qüestió. E. coli pot viure alimentant-se de dos tipus de sucre molt diferents: la glucosa i la lactosa. Alimentat únicament amb qualsevol dels dos sucres, el bacteri comença a dividir-se ràpidament, amb un temps de duplicació d’uns vint minuts. La taxa de creixement es pot representar en forma de corba exponencial —u, dos, quatre, vuit, setze…—, que augmenta fins que el cultiu es torna tèrbol i la font de sucre s’exhaureix.
Aquesta corba de creixement imparable va deixar captivat el biòleg francès Jacques Monod. Monod havia tornat a París el 1937 després d’haver passat un any estudiant mosques amb Thomas Morgan al California Institute of Technology. L’estada de Monod a Califòrnia no havia sigut gaire fructífera —s’havia passat la major part del temps tocant Bach amb l’orquestra local i aprenent dixie i jazz—, però París era una ciutat assetjada i opressiva. L’estiu del 1940, Bèlgica i Polònia ja havien caigut en mans dels alemanys. Pel juny d’aquell any, França, que havia tingut moltíssimes baixes en combat, va firmar un armistici que permetia a l’exèrcit alemany ocupar bona part del nord i l’oest del país.
París va ser declarada «ciutat oberta», lliure de bombardejos i de devastació, però accessible a les tropes nazis. Es van evacuar els infants, els museus es van buidar de quadres i es van tapar els aparadors amb fustes. «París serà sempre París», cantava Maurice Chevalier l’any 1939, encara que fos com en un prec; però la «ciutat de la llum» estava ben poc il·luminada. Els carrers eren sinistres; els cafès eren buits; a la nit hi havia apagades periòdiques que la submergien en una foscor anguniosament inhòspita.
A la tardor del 1940, mentre a tots els edificis oficials onejaven banderes vermelles i negres amb l’esvàstica i les tropes alemanyes anunciaven amb altaveus pels Camps Elisis el toc de queda nocturn, Monod treballava amb E. coli en unes golfes caloroses i fosques de la Sorbona (aquell mateix any es va unir a la resistència francesa, però molts dels seus col·legues no van arribar a tenir coneixement de les seves activitats polítiques). A l’hivern, en un laboratori quasi glaçat per la gelor de fora on havia de matar el temps fins a migdia —esperant que se li desglacés una mica d’àcid acètic i complint la penitència d’haver d’escoltar al carrer la propaganda nazi—, Monod va repetir l’experiment de cultiu bacterià, però fent-hi una modificació estratègica. Aquesta vegada va fer el cultiu posant-hi junts glucosa i lactosa, dos sucres diferents.
Si tots els sucres eren sucre —és a dir, si el metabolisme de la lactosa no era diferent del de la glucosa—, hauria sigut d’esperar que els bacteris s’haguessin alimentat de la barreja de glucosa i lactosa seguint la mateixa corba suau de creixement. Monod, però, va trobar una distorsió en els resultats; una distorsió en sentit literal. Al començament, d’acord amb les previsions, les bacteris s’havien multiplicat de forma exponencial, però de cop el creixement s’havia aturat per reprendre al cap d’una mica. Monod va investigar el perquè d’aquella pausa, i va descobrir un fenomen inusual. Les cèl·lules d’E. coli no havien consumit tots dos sucres indistintament, sinó selectivament, alimentant-se primer de la glucosa. A continuació els bacteris havien parat de multiplicar-se, com si es plantegessin un possible canvi de dieta, havien passat a alimentar-se de lactosa i havien continuat multiplicant-se. Monod va donar a aquest fenomen el nom de diàuxia (‘creixement en dues fases’).
Aquesta inflexió en la corba de creixement, tot i ser petita, va desconcertar Monod. Li feia nosa, com una brossa a l’ull del seu instint de científic. Un bacteri que s’alimentés de sucres hauria hagut de multiplicar-se seguint una corba suau. ¿Per què el canvi de consum de sucres causava una interrupció en el creixement? ¿Com podia un bacteri «saber», o ni tan sols percebre, que la font de sucre havia canviat? I ¿per què consumia un sucre primer, i el segon només quan el primer s’havia acabat, igual que un dinar de restaurant de dos plats?
A finals de la dècada del 1940, Monod havia descobert que la distorsió era conseqüència d’una adaptació metabòlica. Quan els bacteris passaven de consumir glucosa a consumir lactosa, produïen enzims de digestió específica de la lactosa; i, quan tornaven a consumir glucosa, aquests enzims desapareixien i tornaven a produir-se enzims de digestió de la glucosa. La producció d’aquests enzims durant el canvi —com un canvi de coberteria entre dos plats d’un àpat (retirar el ganivet del peix i posar la forquilla de les postres)— exigia uns quants minuts, cosa que explicava la interrupció que s’observava en el creixement.
Segons Monod, la diàuxia portava a pensar que els gens es podien regular per mitjà d’estímuls metabòlics. L’aparició o desaparició dels enzims a la cèl·lula —que són proteïnes, i per tant són codificades pels gens— havia ser ser induïda pels gens, que devien actuar com interruptors moleculars que s’activen o es desactiven. A principis de la dècada del 1950, a París, Monod, juntament amb François Jacob, va començar a estudiar de manera sistemàtica la regulació gènica d’E. coli per mitjà de mutacions provocades, el mètode que Morgan havia utilitzat amb tant d’èxit amb les mosques del vinagre.[032]
Igual que en el cas de les mosques, els bacteris mutants van proporcionar resultats reveladors. Monod i Jacob, amb la col·laboració d’Arthur Pardee, un genetista microbià dels Estats Units, van descobrir tres principis cardinals que governaven la regulació gènica. En primer lloc, quan un gen s’activava o es desactivava, l’ADN original es conservava intacte a l’interior de la cèl·lula. La feina la realitzava l’ARN: quan un gen s’activava, era induït a fer més ARN missatger i per tant a produir més enzims de digestió del sucre. La identitat metabòlica d’una cèl·lula —és a dir, si era consumidora de lactosa o de glucosa— es podia conèixer no per la seva seqüència gènica, que sempre era la mateixa, sinó per la quantitat d’ARN que produïa un gen concret. Durant el metabolisme de la lactosa, els ARN dels enzims de digestió de la lactosa eren abundants; durant el metabolisme de la glucosa, en canvi, aquests missatgers es reprimien, i els ARN que abundaven eren els dels enzims de digestió de la glucosa.
En segon lloc, la producció d’ARN missatger es regulava de forma coordinada. Quan la font de sucre passava a ser la lactosa, els bacteris activaven tot un grup de gens —diferents gens metabolitzadors de la lactosa— per digerir la lactosa: un dels gens del grup codificava una «proteïna transportadora» que permetia a la lactosa accedir a l’interior de la cèl·lula bacteriana; un altre gen codificava un enzim necessari per escindir la lactosa en dues parts; i encara n’hi havia un altre que codificava un enzim responsable de degradar aquestes dues parts en substàncies químiques més petites. Cosa curiosa, tots els gens responsables d’una via metabòlica concreta estaven físicament l’un al costat de l’altre al cromosoma bacterià —igual que els llibres d’un mateix tema en una biblioteca—, i a la cèl·lula eren induïts de forma simultània. L’alteració metabòlica produïa a la cèl·lula una profunda alteració genètica. No era un canvi de coberteria i prou; era tot el servei de taula, el que es canviava de cop. Tota una cadena funcional de gens s’activava i es desactivava com si la regulés un gestor de tasques comú o un interruptor general. Monod va donar a aquest tipus d’unitats genètiques el nom d’operó.[033]
La síntesi de proteïnes, doncs, anava perfectament sincronitzada amb els requeriments del medi: si s’hi aportava un sucre o un altre, s’activava tot el grup de gens corresponent per metabolitzar-lo. La formidable economia evolutiva havia trobat una altra solució elegantíssima per a la regulació gènica. Sense el gen no hi havia missatge, i així no hi havia cap proteïna que treballés perquè sí.
* * *
¿Què feia que una proteïna sensible a la lactosa reconegués i regulés únicament el gen de digestió de la lactosa, però no els milers d’altres gens de la cèl·lula? Monod i Jacob van descobrir que la tercera característica cardinal de la regulació gènica era que cada gen tenia incorporada una seqüència específica d’ADN amb funció reguladora que actuava com una etiqueta identificativa. Quan una proteïna sensible a un sucre detectava aquest sucre al medi, identificava l’etiqueta i feia activar el gen corresponent. Això feia que el gen fabriqués més ARN missatgers i que, per tant, es produís l’enzim apropiat per digerir el sucre.
Un gen, en definitiva, contenia no només la informació per sintetitzar una proteïna sinó també la informació sobre quan i on havia de fabricar-la. Totes aquestes dades estaven codificades en l’ADN, normalment incorporades a l’inici de cada gen (tot i que aquestes seqüències de funció reguladora també poden estar situades al final o a la meitat). La combinació de les seqüències reguladores i la seqüència codificadora de la proteïna constituïen un gen.
Hem de tornar una vegada més a l’analogia que més amunt hem fet amb el llenguatge humà. Quan l’any 1910 Morgan va descobrir el lligament genètic, no havia vist cap lògica aparent en el fet que un gen estigués lligat físicament amb un altre en el cromosoma: semblava que el gen del color de cos negrós (sable) i el gen dels ulls blancs no tinguessin cap mena de vincle funcional, però estaven units com carn i ungla en un mateix cromosoma. Al model de Jacob i Monod, en canvi, els gens bacterians estaven units amb una finalitat. Els gens relacionats amb una mateixa via metabòlica estaven lligats físicament entre ells: qui convivia fent una mateixa feina, convivia al genoma. Hi havia unes seqüències específiques d’ADN que anaven unides a un gen per posar-lo en situació respecte a l’activitat —la «feina»— que havia de fer. Aquestes seqüències, destinades a activar o desactivar el gen, es podrien comparar amb notes o signes de puntuació —cometes, una coma, una inicial majúscula— d’una frase: donen context, èmfasi i sentit, informen el lector de quines parts s’han de llegir juntes i quan ha de parar perquè comença la frase següent:
«Així és l’estructura del nostre genoma. Conté, entre altres coses, unitats que es regulen de forma independent. Hi ha paraules ajuntades en forma de frases, mentre que d’altres estan separades per punts i comes, comes, i punts».
Pardee, Jacob i Monod van publicar el seu monumental estudi sobre l’operó de la lactosa el 1959, sis anys després de l’assaig de Watson i Crick sobre l’estructura de l’ADN. Batejat amb el nom d’estudi Pa-Ja-Mo a partir de les dues primeres lletres de tots tres autors —col·loquialment Pajama, «pijama» en anglès—, es va convertir immediatament en un clàssic que ha tingut grans repercussions en biologia. Els gens, segons l’estudi Pajama, no eren simples plantilles passives. Encara que totes les cèl·lules comparteixen un mateix conjunt de gens —un genoma idèntic—, l’activació o repressió selectiva de subconjunts de gens concrets permet a una cèl·lula determinada reaccionar en funció del medi. El genoma és una plantilla activa, capaç de fer ús de parts determinades del seu codi en moments i circumstàncies diverses.
Les proteïnes actuen en aquest procés com sensors reguladors o interruptors generals, activant i desactivant de forma coordinada gens o combinacions de gens. Igual que la partitura general d’una peça simfònica de complexitat descomunal, el genoma conté les instruccions per al desenvolupament i la preservació dels organismes. Aquesta «partitura» genòmica, però, és inactiva sense les proteïnes. Les proteïnes fan realitat aquesta informació activant o reprimint uns gens o uns altres. (Algunes d’aquestes proteïnes reguladores també reben el nom de factors de transcripció). «Dirigeixen» el genoma i n’interpreten la música: activen la viola en el minut catorze, un cop de címbal durant l’arpegi, un redoblament de timbales al crescendo. El concepte seria:
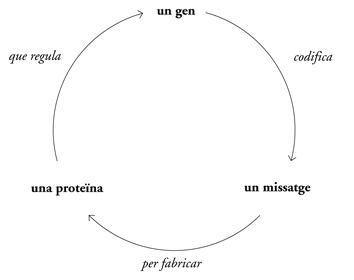
L’estudi Pa-Ja-Mo liquidava una qüestió cabdal de la genètica: ¿com pot ser que un organisme tingui un conjunt fixat de gens però reaccioni de manera tan precisa als canvis del medi? I també suggeria una solució a una qüestió d’embriogènesi igualment cabdal: ¿com pot ser que d’un embrió en què totes les cèl·lules tenen el mateix conjunt de gens sorgeixin milers de tipus cel·lulars diferents? La regulació dels gens —l’activació i desactivació selectiva de determinats gens a determinades cèl·lules i en moments determinats— havia d’aportar un grau essencial de complexitat a la naturalesa impertorbable de la informació biològica.
Monod sostenia que era gràcies a la regulació gènica que les cèl·lules podien desenvolupar les seves funcions específiques en el temps i en l’espai. «El genoma conté no només una sèrie de plantilles [és a dir, els gens], sinó també un programa coordinat […] i els mitjans de regular-ne l’execució», acabaven dient Monod i Jacob. Tant els glòbuls vermells com les cèl·lules hepàtiques de Walter Noel contenien la mateixa informació genètica, però la regulació gènica es cuidava de fer que la proteïna de l’hemoglobina fos present només als glòbuls vermells, i al fetge no. L’eruga i la papallona contenen exactament el mateix genoma, però la regulació gènica fa possible que l’una es metamorfosi en l’altra.
L’embriogènesi es podriar reformular com el descabdellament gradual de la regulació gènica a partir de l’embrió unicel·lular. Aquest seria el «moviment» que Aristòtil s’havia imaginat segles enrere de forma tan viva. Segons una anècdota cèlebre, algú va demanar a un cosmòleg medieval què sostenia la terra.
—Unes tortugues —va dir.
—I ¿què sosté les tortugues? —li va demanar l’altre.
—Més tortugues.
—¿I aquestes tortugues?
—No ho entén —va dir el cosmòleg, picant de peus—. Són tot tortugues!
Per a un genetista, el desenvolupament d’un organisme es podria definir com la inducció (o repressió) seqüencial dels gens i les cadenes genètiques. Els gens codificaven proteïnes que activaven gens que codificaven proteïnes que activaven gens, i així successivament fins a arribar a la primera cèl·lula embriològica. Eren tot gens.[034]
* * *
La regulació gènica —l’activació i desactivació dels gens per obra de les proteïnes— definia el mecanisme per mitjà del qual tota la complexitat combinatòria es podia formar a partir de l’únic document d’informació genètica de la cèl·lula, però no explicava pròpiament el fenomen de còpia dels gens: ¿com es repliquen els gens quan una cèl·lula es divideix en dues, o quan es forma un espermatozou o un òvul?
Als ulls de Watson i Crick, el model d’ADN de doble hèlix —amb dos filaments «yin-yang» complementaris contraposats l’un a l’altre— insinuava el mecanisme de replicació mateix. Al final del seu article del 1953 comentaven: «No ens ha passat desapercebut que l’aparellament específic [d’ADN] que hem postulat suggereix immediatament un possible mecanisme de còpia del material genètic». El seu model d’ADN no era una imatge bonica i prou, sinó que la seva mateixa estructura anticipava les característiques principals de la seva funció. Watson i Crick van postular que cada filament d’ADN s’utilitzava per obtenir una còpia de si mateix, de tal manera que de la doble hèlix originària en sortien dues. Durant la replicació, els filaments yin-yang d’ADN se separaven. El yin servia de plantilla per fer un yang i el yang servia per fer un yin, i així s’obtenien dos parells yin-yang. (Matthew Meselson i Frank Stahl van demostrar l’existència d’aquest mecanisme l’any 1958).
Una doble hèlix d’ADN, però, no és capaç de fer una còpia de si mateixa pel seu compte; si no fos així, es replicaria sense cap mena de control. Era probable que hi hagués un enzim —una proteïna replicadora— que es dediqués a copiar l’ADN. El 1957, el bioquímic Arthur Kornberg es va proposar d’aïllar l’enzim que copiava l’ADN. Kornberg va pensar que, si aquest enzim existia, el lloc més fàcil de trobar-lo seria en un organisme que es dividís de pressa, com ara el bacteri E. coli en la seva fase de creixement més desenfrenada.
El 1958, Kornberg s’havia dedicat a purificar i tornar a purificar el residu bacterià fins a obtenir una preparació d’enzim pràcticament pura («Els genetistes compten, i els bioquímics netegen», em va dir una vegada). Va batejar-lo amb el nom de polimerasa d’ADN (l’ADN és un polímer d’A, C, G i T, i aquest era l’enzim sintetitzador del polímer). Quan va afegir l’enzim purificat a l’ADN, juntament amb una font d’energia i una provisió de bases de nucleòtids —A, T, G i C–, va veure que al tub d’assaig es formaven noves cadenes d’àcid nucleic: l’ADN fet a la seva pròpia imatge.
«Cinc anys enrere», va escriure Kornberg el 1960, «la síntesi de l’ADN era considerada un procés “vital”», una reacció misteriosa que no era possible de reproduir en un tub d’assaig per mera addició o substracció de substàncies químiques. Segons aquesta teoria, «potinejar el material genètic [de la vida] mateix no podia conduir sinó al desordre». La síntesi d’ADN feta per Kornberg, però, havia creat ordre a partir del desordre: s’havia obtingut un gen a partir de les seves subunitats químiques. La inexpugnabilitat dels gens havia deixat de ser una barrera.
Aquí convé fer un petit recordatori: com totes les proteïnes, la polimerasa d’ADN, és a dir l’enzim que permet la replicació de l’ADN, també és producte d’un gen.[035] A cada genoma, per tant, hi ha continguts els codis per fabricar les proteïnes que permetran que el genoma mateix es reprodueixi. Aquest grau addicional de complexitat —el fet que l’ADN codifiqui una proteïna que permet a l’ADN replicar-se— és important perquè proporciona un principi de regulació determinant. La replicació de l’ADN es pot activar o desactivar per mitjà d’altres senyals o reguladors, com per exemple l’edat o l’estat nutricional de la cèl·lula, cosa que permet a les cèl·lules fer còpies d’ADN només quan estan en condicions de dividir-se. Aquest sistema té una possible pega: que, si els reguladors es descontrolen, no hi ha res que pugui impedir a una cèl·lula de replicar-se sense parar. Això, com aviat veurem, és el pitjor mal dels gens defectuosos: el càncer.
* * *
Els gens fan proteïnes que regulen els gens. Els gens fan proteïnes que fan replicar-se els gens. La tercera R de la fisiologia dels gens és una paraula que no pertany al vocabulari del comú de la gent, però és essencial per a la supervivència de la nostra espècie: recombinació, és a dir, la capacitat de produir noves combinacions de gens.
Per entendre què és la recombinació hauríem de tornar una vegada més a Mendel i a Darwin. Un segle de recerca genètica va permetre comprendre com els organismes es transmeten la «semblança» els uns als altres. Les unitats d’informació hereditària, codificades a l’ADN i empaquetades als cromosomes, es transmeten a l’embrió per mitjà de l’espermatozou i l’òvul, i de l’embrió a totes les cèl·lules del cos d’un organisme. Aquestes unitats codifiquen missatges per fabricar les proteïnes; i els missatgers i les proteïnes, per la seva banda, fan realitat la forma i la funció de l’organisme.
Ara bé, tot i que aquesta descripció del mecanisme de l’herència resolia la pregunta de Mendel —¿per què els fills s’assemblen als pares?—, no aconseguia resoldre l’enigma invers de Darwin: ¿per què els fills poden no assemblar-se als pares? Perquè l’evolució tingui lloc, un organisme ha de ser capaç de produir variació genètica, és a dir que ha de tenir una descendència que sigui genèticament diferent de tots dos progenitors. Si en principi els gens transmeten semblança, ¿com s’explica que puguin transmetre «dissemblança»?
Un mecanisme de producció de variació en la naturalesa és la mutació, que consisteix en alteracions en la seqüència de l’ADN (per exemple, una A substituïda per una T) que poden modificar l’estructura d’una proteïna i, per tant, alterar-ne la funció. Les mutacions s’originen quan l’ADN queda danyat per substàncies químiques o pels rajos X, o quan l’enzim de replicació de l’ADN comet un error accidental durant la còpia dels gens. Però també hi ha un altre mecanisme de generació de diversitat genètica, que és la possibilitat que la informació genètica s’intercanviï entre cromosomes. Part de l’ADN del cromosoma matern pot intercanviar-se amb ADN del cromosoma patern, per acabar generant un gen híbrid a partir del gen matern i el gen patern. La recombinació també és una forma de «mutació», amb la diferència que entre els cromosomes s’intercanvien fragments considerables de material genètic.[036]
El pas d’informació genètica d’un cromosoma a un altre té lloc únicament en circumstàncies especialíssimes. Una és quan es produeixen espermatozous i òvuls; poc abans de l’espermatogènesi i l’oogènesi, la cèl·lula es converteix breument en un pati de jugar per als gens. Els cromosomes homòlegs matern i patern s’abracen i intercanvien informació genètica sense cap inconvenient. L’intercanvi d’informació genètica entre cromosomes homòlegs és essencial perquè hi hagi una barreja i una correspondència d’informació hereditària dels progenitors. Morgan va donar a aquest fenomen el nom d’entrecreuament (els seus alumnes havien recorregut a l’entrecreuament per elaborar un mapa dels gens de les mosques). El terme que posteriorment s’ha adoptat és el de recombinació, que consisteix en la capacitat de generar noves combinacions de combinacions de gens.
L’altra circumstància és més extraordinària encara. Quan l’ADN sofreix una alteració per acció d’algun mutagen, com per exemple els rajos X, la informació genètica queda evidentment danyada; i, quan hi ha aquest dany, el gen es pot tornar a copiar a partir de la còpia «bessona» que hi ha en el cromosoma homòleg: part de la còpia materna es pot reescriure a partir de la còpia paterna (i viceversa), cosa que de retruc té com a resultat la formació de gens híbrids.
També en aquest cas l’aparellament de bases s’utilitza per reconstruir el gen. El yin repara el yang, la còpia restitueix l’original: amb l’ADN, igual que amb el Dorian Gray d’Oscar Wilde, el prototip rep constantment forces renovades del seu propi retrat. Les proteïnes dirigeixen i coordinen tot el procés —conduint el filament danyat cap al gen intacte, copiant i corregint la informació perduda i cosint els punts de trencament— i en definitiva fan possible la transferència d’informació del filament intacte al filament danyat.
* * *
Regulació. Replicació. Recombinació. Convé tenir present que aquestes tres R de la fisiologia del gen depenen profundament de l’estructura molecular de l’ADN, és a dir, de l’aparellament de bases de la doble hèlix proposat per Watson i Crick.
La regulació gènica té lloc per mitjà de la transcripció d’ADN a ARN, que està en funció de la seva estructura de parells de bases. Quan una cadena d’ADN s’utilitza per sintetitzar l’ARN missatger, és l’aparellament de bases entre l’ADN i l’ARN el que permet que un gen es copiï en forma d’ARN. En el cas de la replicació, l’ADN també és copiat fent-se servir ell mateix com a referència. Cada cadena serveix per sintetitzar una versió complementària de si mateixa, cosa que dóna com a resultat una doble hèlix que s’escindeix en dues dobles hèlixs. D’altra banda, durant la recombinació de l’ADN l’estratègia d’acarar base contra base es torna a utilitzar per restituir l’ADN danyat. La còpia danyada d’un gen es reconstrueix recorrent a la cadena complementària, amb una altra còpia del gen, com a referència.[037]
La doble hèlix ha resolt els tres reptes principals de la fisiologia genètica, servint-se d’enginyoses variacions sobre el mateix tema. S’utilitzen substàncies químiques que fan d’imatge especular per sintetitzar substàncies químiques especulars, és a dir reflexos per reconstruir la imatge original; parelles per mantenir la fidelitat i la inamobilitat de la informació. «Monet és un ull i prou», va dir Cézanne del seu amic una vegada; «però quin ull, Déu meu!». Seguint el mateix fil de pensament es pot dir: l’ADN és una substància química i prou; però quina substància química, Déu meu!
* * *
En biologia existeix una distinció tradicional entre dos grups de científics: els anatomistes i els fisiòlegs. Els anatomistes descriuen la naturalesa dels elements materials, les estructures i les parts del cos; descriuen com són les coses. Els fisiòlegs, per la seva banda, estudien els mecanismes per mitjà dels quals aquestes estructures i parts es relacionen entre elles per fer possibles les funcions dels éssers vius; és a dir, s’interessen per com funcionen les coses.
Aquesta distinció assenyala també una transició cabdal en la història de la genètica. Mendel va ser segurament el primer «anatomista» del gen: revelant la transmissió d’informació al llarg de generacions de pesoleres, havia definit l’estructura íntima del gen com un corpuscle indivisible d’informació. Morgan i Sturtevant havien prolongat aquesta línia anatomista a la dècada del 1920 demostrant que els gens eren unitats materials que es disposaven linealment al llarg dels cromosomes. Entre les dècades del 1940 i el 1950, Avery, Watson i Crick van identificar l’ADN com la molècula del gen i en van descriure l’estructura com una doble hèlix, amb la qual cosa van portar la noció anatòmica del gen a la seva culminació natural.
Entre finals dels anys cinquanta i els anys setanta, però, va ser la fisiologia dels gens el que va dominar la recerca científica. El fet que els gens es poguessin regular —és a dir, que hi hagués uns estímuls concrets que els poguessin activar o desactivar— va ajudar a comprendre com actuen els gens en el temps i en l’espai per determinar els trets peculiars dels diferents tipus de cèl·lules; i el fet que poguessin reproduir-se, recombinar-se entre cromosomes i ser reparats per unes proteïnes específiques va explicar de quina manera les cèl·lules i els organismes aconsegueixen conservar, copiar i reconfigurar la informació genètica a través de les generacions.
Per als especialistes en biologia humana, cada un d’aquests descobriments va comportar uns beneficis molt importants. Amb el pas que en genètica es va fer d’una noció materialista del gen a una idea mecanicista —de què són els gens a què fan—, els biòlegs van començar a observar relacions llargament buscades entre els gens, la fisiologia humana i la patologia. Una malaltia podia aparèixer no només per una alteració en el codi genètic corresponent a una proteïna (com per exemple l’hemoglobina en el cas de l’anèmia falciforme), sinó també a causa de la regulació gènica (per la incapacitat d’activar o desactivar el gen adequat en la cèl·lula adequada i el moment adequat). La replicació genètica havia d’explicar per què un organisme pluricel·lular sorgia d’una única cèl·lula, i els errors en la replicació havien d’aclarir per què una malaltia metabòlica d’aparició sobtada o un trastorn mental greu podien aparèixer en una família on no n’hi havia hagut cap cas fins aleshores. Les similituds entre els genomes havien d’explicar les semblances entre pares i fills, i les mutacions i la recombinació havien d’explicar-ne les diferències. Les famílies no compartien només xarxes socials i culturals, sinó també xarxes de gens dinàmics.
De la mateixa manera que l’anatomia humana del segle XIX va posar els fonaments per a la medicina del segle XX, l’anatomia i la fisiologia dels gens posaran els fonaments d’una ciència biològica nova i plena de força. Al llarg de les dècades següents, aquesta ciència revolucionària eixamplarà el seu camp d’estudi des dels organismes simples fins als organismes complexos. El seu vocabulari conceptual —regulació gènica, recombinació, mutació, reparació de l’ADN— saltarà de les revistes purament científiques als llibres de text mèdics, i acabarà introduint-se en el debat social i cultural (el terme raça, com veurem tot seguit, no es pot comprendre plenament sense tenir abans una comprensió del que són la recombinació i la mutació). La nova ciència intentarà explicar de quina manera els gens fan que els éssers humans es formin, es conservin, es refacin i es reprodueixin, i també de quina manera les variacions en l’estructura i la fisiologia dels gens poden influir en les variacions observades en la identitat, el caràcter, la salut i les malalties dels éssers humans.