KAPITEL 7
Äpfel oder Indianer
Warum es den Bewohnern mancher Regionen nicht gelang, Pflanzen zu domestizieren
Wir haben gerade erfahren, wie die Völker einiger Regionen begannen, Wildpflanzen zu kultivieren, und damit einen Schritt mit ebenso schwerwiegenden wie unabsehbaren Folgen für ihr Leben und den Platz ihrer Nachkommen in der Geschichte taten. Kehren wir nun zurück zu der Frage, warum die Landwirtschaft in einigen fruchtbaren, höchst geeigneten Regionen wie Kalifornien, Europa, Teilen Australiens mit gemäßigtem Klima und Afrika südlich des Äquators nicht unabhängig entstand, und weiter, warum sie in einigen der unabhängigen Entstehungszentren sehr viel früher in Erscheinung trat als in anderen.
Als Antwort bieten sich zwei gegensätzliche Erklärungen an: Schuld war entweder die örtliche Bevölkerung oder das örtliche Angebot an Wildpflanzen. Die eine Erklärung geht davon aus, daß in allen ausreichend bewässerten Gebieten der gemäßigten oder tropischen Breiten eine genügend große Zahl von Wildpflanzen, die sich zur Domestikation eignen, vorhanden sind. In diesem Fall wäre die Erklärung für die Nichtentstehung der Landwirtschaft in einigen dieser Gebiete in kulturellen Eigenschaften ihrer Bewohner zu suchen. Die andere Erklärung unterstellt, daß in jeder der großen Weltregionen wenigstens einige Menschen die Experimentierfreudigkeit besessen hätten, die zur Domestikation führte. Nur ein Fehlen geeigneter Pflanzenarten könnte dann erklären, warum die Landwirtschaft in einigen Gebieten nicht auf den Plan trat.
Wie ich im nächsten Kapitel zeigen werde, ist das entsprechende Problem im Fall der Domestikation großer Säugetiere sehr viel einfacher zu lösen, was an der sehr geringen Zahl von Säugetier- im Vergleich zu Pflanzenarten liegt. Auf der ganzen Welt gibt es nämlich nur etwa 148 Arten großer, wildlebender Landsäugetiere, die als Domestikationskandidaten prinzipiell in Betracht kommen. Die Eignung einzelner Arten hängt von einer recht begrenzten Zahl von Faktoren ab. Deshalb kann für eine bestimmte Region ohne weiteres geprüft werden, ob die Nichtdomestikation dortiger Kandidaten womöglich auf das Fehlen geeigneter Arten anstatt auf irgendwelche Besonderheiten der örtlichen Vertreter der Spezies Mensch zurückzuführen ist.
Ein solches Vorgehen würde bei Pflanzen schon an der riesigen Artenzahl scheitern, gibt es doch nicht weniger als 200 000 Arten von Blütenpflanzen, also jener in der Vegetation überwiegenden Gewächse, von denen die meisten unserer Kulturpflanzen abstammen. Es wäre ein aussichtsloses Unterfangen, alle Wildpflanzen selbst eines so begrenzten Gebiets wie Kalifornien untersuchen zu wollen, um zu beurteilen, wie viele von ihnen domestizierbar gewesen wären. Wir werden aber gleich sehen, wie sich dieses Problem umgehen läßt.
Angesichts der gewaltigen Zahl von Blütenpflanzen könnte einem zunächst der Gedanke kommen, daß bei solcher Überfülle jede Region mit einigermaßen mildem Klima mehr als genügend Arten aufweisen müßte, um daraus eine Vielzahl von Kulturgewächsen zu züchten.
Man darf aber nicht vergessen, daß die allermeisten Wildpflanzen aus offenkundigen Gründen als Nahrung nicht in Betracht kommen: Sie sind holzig, tragen keine eßbaren Früchte, und ihre Blätter und Wurzeln sind ebenfalls nicht genießbar. Von den rund 200 000 Wildpflanzenarten stehen nur wenige Tausend auf dem Speiseplan des Men schen, und nur einige Hundert davon wurden mehr oder weniger domestiziert. Selbst von dieser relativ kleinen Zahl von Kulturpflanzen tragen die meisten nur unwesentlich zu unserer Ernährung bei und hätten allein kein ausreichendes Fundament für die Errichtung von Zivilisationen abgegeben. Über 80 Prozent der pflanzlichen Nahrung, die heute jährlich erzeugt wird, entfällt auf ein bloßes Dutzend Arten. Dazu zählen Weizen, Mais, Reis, Gerste und Sorghum (Getreide), Sojabohnen (Hülsenfrüchte), Kartoffeln, Maniok und Süßkartoffeln (Wurzelknollen), Zuckerrohr, Zuckerrüben (Zuckerpflanzen) und Bananen (Obst). Über die Hälfte des Kalorienverbrauchs der Weltbevölkerung wird heute allein durch Getreide gedeckt. In Anbetracht einer so geringen Zahl bedeutender Kulturpflanzen, die ausnahmslos schon vor Jahrtausenden domestiziert wurden, mag es nicht mehr ganz so überraschen, daß in vielen Regionen der Welt keine Wildpflanzen mit derart überragendem Potential als Nahrungspflanzen heimisch waren. Daß in der Neuzeit keine einzige bedeutende Anbaupflanze domestiziert wurde, legt den Schluß nahe, daß in vorgeschichtlicher Zeit schon praktisch sämtliche für den Menschen nützlichen Wildpflanzen erkundet und domestiziert wurden, sofern das eben möglich und lohnend war.
Dennoch gibt es einige Fälle, in denen schwer zu erklären ist, warum bestimmte Wildpflanzen nicht domestiziert wurden. Am auffälligsten sind jene, wo bestimmte Pflanzen in einem Gebiet domestiziert wurden, in einem anderen jedoch nicht. In diesen Fällen steht außer Zweifel, daß es möglich war, aus den betreffenden Wildpflanzen nützliche Anbaugewächse zu züchten, so daß wir fragen müssen, warum die Domestikation in manchen Gebieten nicht stattfand.
Ein solches Rätsel gibt uns Afrika auf. Sorghum, eine wichtige Getreidepflanze, wurde in der Sahelzone südlich der Sahara domestiziert. Die Wildform kommt auch weiter südlich vor und ist selbst im südlichen Afrika heimisch, wo jedoch weder Sorghum noch irgendeine andere Pflanze kultiviert wurde, bis vor 2000 Jahren die Bantu-Bauern aus den Gebieten nördlich des Äquators ihr komplettes Bündel von Anbaupflanzen mitbrachten. Warum, so fragt man sich, kamen die Völker des südlichen Afrika nicht von selbst auf die Idee, Sorghum zu domestizieren?
Nicht minder rätselhaft ist das Ausbleiben der Domestikation von Flachs in seinem natürlichen Verbreitungsgebiet in Westeuropa und Nordafrika oder von Einkornweizen in seinem Verbreitungsgebiet im südlichen Balkan. Da beide Pflanzen unter den ersten acht Kulturpflanzen des Fruchtbaren Halbmonds waren, zählten sie vermutlich zu den besonders leicht domestizierbaren Wildpflanzen. Außerhalb Vorderasiens wurden sie als Anbaupflanzen erst übernommen, als sie zusammen mit dem gesamten Ensemble landwirtschaftlicher Techniken aus Vorderasien eintrafen. Warum hatten die Völker dieser Regionen mit ihrem Anbau nicht schon früher aus eigenem Antrieb begonnen?
Ein ähnliches Beispiel betrifft die vier ältesten domestizierten Obstgewächse Vorderasiens, deren natürliche Verbreitungsgebiete weit über den östlichen Mittelmeerraum, wo sie dem Anschein nach zuerst domestiziert wurden, hinausreichten: Oliven, Weinbeeren und Feigen waren auch in Italien, Spanien und Nordwestafrika heimisch, während die Dattelpalme in ganz Nordafrika und Arabien wuchs. Diese vier zählten offensichtlich zu den am leichtesten domestizierbaren Fruchtbäumen. Warum wurden sie außerhalb Vorderasiens nicht ebenfalls domestiziert, sondern erst angebaut, nachdem sie im östlichen Mittelmeerraum domestiziert und von dort als fertige Kulturgewächse importiert worden waren?
Weitere krasse Beispiele sind Wildpflanzen, die in Gebieten, in denen sich die Landwirtschaft nicht von selbst entwickelte, nie domestiziert wurden, obwohl aus engen Verwandten dieser Arten anderswo erfolgreich Kulturpflanzen gezüchtet wurden. So wurde beispielsweise im östlichen Mittelmeerraum die Olivenart Olea europea domestiziert. Im tropischen und südlichen Afrika, in Südasien und Ostaustralien kommen etwa 40 andere Olivenarten vor, die zum Teil eng mit der Olea europea verwandt sind, von denen jedoch keine einzige jemals domestiziert wurde. Ein ähnliches Beispiel handelt von Äpfeln und Weinbeeren. In Eurasien wurde je eine Apfel- und eine Weinbeerenart domestiziert. Zahlreiche verwandte Apfel- und Weinbeerenarten kommen in Nordamerika vor, und einige von ihnen wurden auch in jüngerer Vergangenheit mit ihren domestizierten eurasischen Pendants gekreuzt, um wertvollere Sorten zu gewinnen. Warum aber hatten die nordamerikanischen Indianer diese doch offenbar nützlichen Gewächse nicht selbst domestiziert?
Die Reihe derartiger Beispiele ließe sich endlos fortsetzen. Die Sache hat aber einen entscheidenden Haken: Man darf sich die Pflanzendomestikation nicht so vorstellen, daß Jäger und Sammler eine einzige Pflanze domestizierten und ansonsten ihr Nomadenleben fortsetzten. Nehmen wir einmal an, aus den nordamerikanischen Wildäpfeln wäre eine erstklassige Kulturpflanze geworden, wenn die örtlichen Jäger und Sammler seßhaft geworden wären und Apfelbäume kultiviert hätten. Aber nomadische Jäger und Sammler hätten ihre bisherige Lebensweise wohl kaum aufgegeben, um sich in Dörfern niederzulassen und Apfelgärten zu bestellen, wenn nicht eine ganze Reihe weiterer domestizierbarer Wildpflanzen und -tiere verfügbar gewesen wären.
Erst dann hätte eine seßhafte bäuerliche Lebensweise womöglich Vorteile gegenüber der Jagd- und Sammelwirtschaft gehabt.
Kurzum, wie beurteilt man das Domestikationspotential der gesamten Flora einer Region? Im Fall der Indianer, die keine Äpfel domestizierten, lautet die Frage: Lag das Problem bei ihnen oder vielmehr bei den Äpfeln?
Um dieser Frage nachzugehen, wollen wir nun drei Regionen vergleichen, die unter den Zentren unabhängiger Domestikation geographisch am weitesten auseinanderliegen. Wie wir sahen, war eine von ihnen, der Bereich des Fruchtbaren Halbmonds, wahrscheinlich das früheste Zentrum der Landwirtschaft und Ursprungsort einiger der wichtigsten Anbaupflanzen der Gegenwart sowie fast sämtlicher wichtiger Haustiere. In Neuguinea und im Osten der heutigen USA, den beiden anderen Regionen, wurden zwar örtlich vorkommende Pflanzen domestiziert, doch ihre Zahl war sehr gering, und nur eine von ihnen erlangte weltweite Bedeutung, so daß das daraus geschnürte Nahrungspaket nicht als Grundlage einer umfassenden technischen und politischen Entwicklung wie in Vorderasien taugte. Im Licht dieses Vergleichs werden wir fragen, ob Pflanzenwelt und Umwelt in Vorderasien eindeutige Vorteile gegenüber Neuguinea und dem Osten der USA besaßen.
Einer der zentralen Tatbestände der Menschheitsgeschichte ist die frühe Bedeutung eines Teils Vorderasiens, der wegen des sichelförmigen Verlaufs seiner Gebirgszüge den Namen »Fruchtbarer Halbmond« trägt (Abbildung 7.1). Diese Region war offenbar die älteste Stätte einer ganzen Kette von Entwicklungen; so entstanden dort Städte, die Schrift, Reiche und das, was wir (im negativen oder positiven Sinne) »Zivilisation« nennen. Voraussetzung all dieser Entwicklungen waren große Siedlungsdichte, die Erwirtschaftung und Speicherung von Nahrungsüberschüssen und das »Durchfüttern« von nicht in der Landwirtschaft tätigen Spezialisten, ermöglicht durch das Aufkommen von Ackerbau und Viehzucht. Die Landwirtschaft war die erste dieser bedeutenden Neuerungen in Vorderasien. Deshalb muß jeder Versuch, die Ursprünge der modernen Welt zu verstehen, auch Antwort auf die Frage geben, warum die in Vorderasien domestizierten Pflanzen und Tiere dieser Region zu einem so gewaltigen Entwicklungsvorsprung verhalfen.
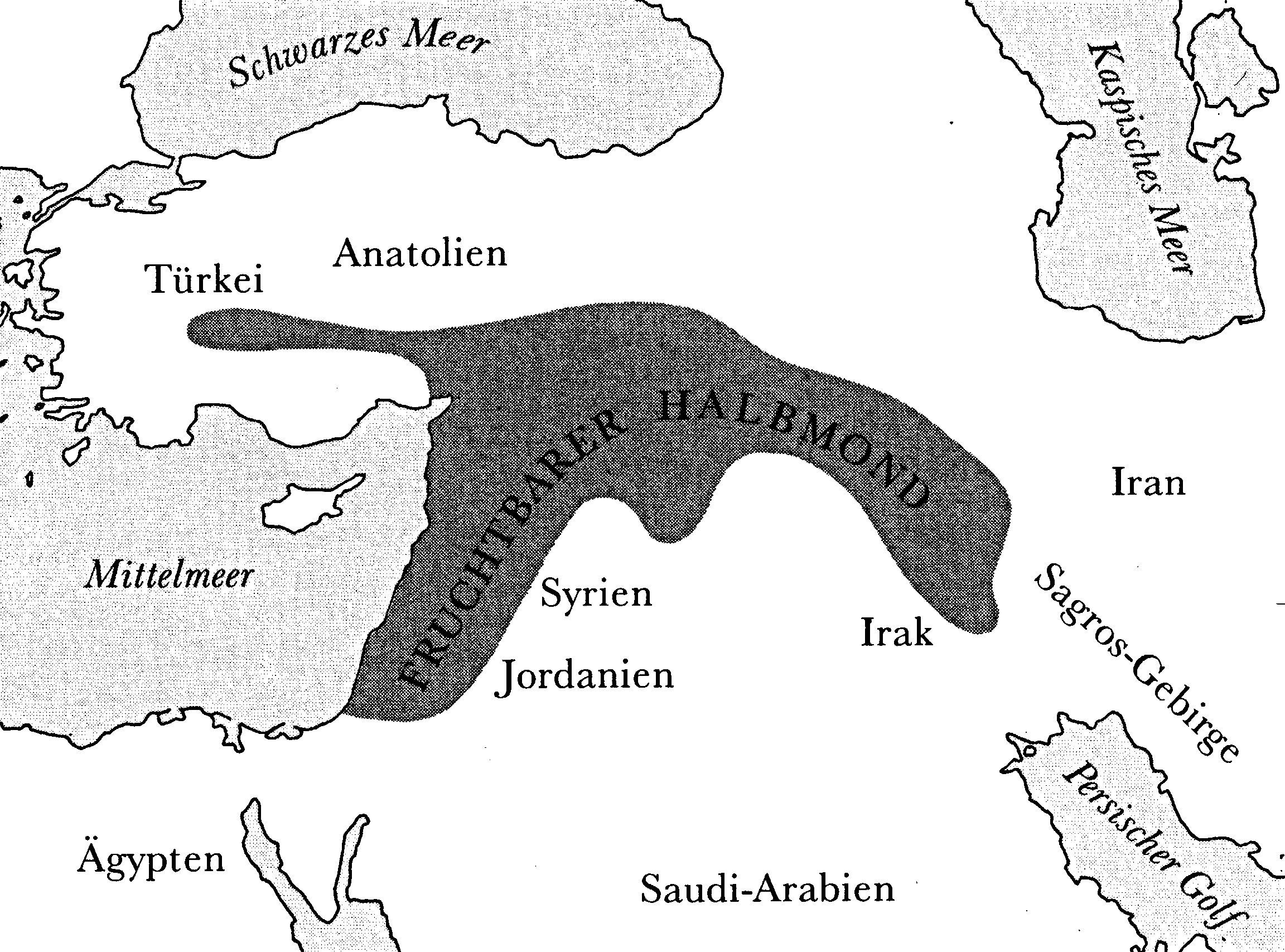
Abbildung 7.1 Fruchtbarer Halbmond – der Teil Vorderasiens, in dem in verschiedenen Gebieten schon vor 7 000 v. Chr. Land wirtschaft betrieben wurde
Glücklicherweise ist die Entstehung der Landwirtschaft im Bereich des Fruchtbaren Halbmonds unter allen Regionen der Erde mit Abstand am gründlichsten untersucht worden. Für die meisten Anbaupflanzen, die dort oder in benachbarten Gebieten domestiziert wurden, konnten die wildwachsenden Vorfahren ermittelt werden; ihre enge Verwandtschaft wurde durch genetische Untersuchungen nachgewiesen; ihr natürliches Verbreitungsgebiet ist bekannt; ihre Veränderungen im Zuge der Domestikation wurden bestimmt und konnten oft einzelnen Genen zugeordnet werden; die Veränderungen sind in übereinanderliegenden Schichten von Ausgrabungsstätten zu beobachten; und schließlich sind auch in etwa Ort und Zeitpunkt der Domestikation bekannt. Ich will nicht abstreiten, daß andere Gebiete, vor allem China, ebenfalls Vorteile als Stätte früher Domestikation besaßen. Am detailliertesten belegt sind diese Vorteile und die aus ihnen resultierende Entstehung von Anbaupflanzen jedoch für Vorderasien.
Einer der Vorteile des Fruchtbaren Halbmonds ist seine Lage inmitten einer mediterranen Klimazone, sprich einem Gebiet mit milden, feuchten Wintern und langen, heißen, trockenen Sommern. Unter solchen klimatischen Bedingungen gedeihen vor allem Pflanzen, die in der Lage sind, die lange Trockenzeit zu überdauern und bei Wiederkehr des Regens schnell aus der Erde zu sprießen. Viele Pflanzen Vorderasiens, insbesondere Gräser und Hülsenfrüchte, paßten sich an diese Bedingungen in einer für den Menschen nützlichen Weise an: Es waren einjährige Pflanzen, die in der Trockenzeit austrocknen und absterben.
Innerhalb ihrer auf ein Jahr beschränkten Lebensspanne bleiben einjährige Pflanzen zwangsläufig kleinwüchsig. Viele investieren ihre Energie statt in hochschießendes Wachstum in die Ausbildung großer Samenkörner, die während der Trockenzeit im Ruhezustand verharren und bei einsetzendem Regen zu sprießen beginnen. Einjährige Pflanzen verwenden mithin, anders als Bäume und Büsche, wenig Energie auf die Erzeugung von Holz und faserigen Stengeln. Dagegen sind viele der großen Samenkörner, insbesondere von einjährigen Getreidearten und Hülsenfrüchten, für Menschen genießbar. Sie stellen sechs der zwölf wichtigsten Anbaupflanzen der Gegenwart dar. Wer indes in waldreicher Umgebung lebt, wird beim Blick aus dem Fenster bemerken, daß die meisten Pflanzenarten vor seinen Augen Bäume und Sträucher sind, die nur einen geringen Teil ihrer Energie in eßbare Samen umwandeln. Zwar gibt es in Gebieten mit feuchtem Klima durchaus Bäume, die große, eßbare Samen hervorbringen, doch diese besitzen nicht die nötige Anpassung, um lange Trockenzeiten unbeschadet zu überstehen, und sind deshalb auch für eine längere Aufbewahrung durch den Menschen ungeeignet.
Ein zweiter Vorteil, den die Flora Vorderasiens gegenüber anderen Regionen besaß, lag darin, daß die wildwachsenden Vorfahren vieler dortiger Kulturpflanzen bereits weit verbreitet und sehr ertragreich waren, so daß ihr Nutzen den örtlichen Sammlern kaum verborgen bleiben konnte. In experimentellen Studien, bei denen Botaniker etwa so, wie es Sammler vor über 10 000 Jahren getan haben mögen, Samen von natürlichem Wildgetreide ernteten, konnte gezeigt werden, daß jährliche Erträge von bis zu einer Tonne Samen pro Hektar möglich sind – das entspricht 50 Kilokalorien Nahrungsenergie bei einem Energieeinsatz von nur einer Kilokalorie. Indem sie große Mengen Wildgetreide innerhalb eines kurzen Zeitraums nach der Samenreife ernteten und anschließend als Nahrungsreserve für den Rest des Jahres lagerten, wurden einige Jäger- und Sammlervölker Vorderasiens schon zu seßhaften Dorfbewohnern, noch bevor sie damit begannen, selbst Pflanzen zu kultivieren.
Da die Getreidearten Vorderasiens schon in ihren Wildformen ertragreich waren, bedurfte es zu ihrer Domestikation nur geringfügiger Veränderungen. Wie im vorigen Kapitel erörtert, stellten sich die wichtigsten davon – der Verlust der natürlichen Samenverbreitungsmittel und des Keimverzugs – automatisch und binnen kurzer Zeit ein, nachdem Menschen damit begonnen hatten, die Samen auf Feldern zu säen. Die wildwachsenden Vorfahren von Weizen und Gerste sehen unseren heutigen Getreidesorten so ähnlich, daß nie Zweifel hinsichtlich der Abstammung aufkamen. Da ihre Domestikation so einfach war, befanden sich großsamige einjährige Pflanzen unter den ersten Anbaugewächsen, die nicht nur in Vorderasien, sondern auch in China und in der Sahelzone Bedeutung erlangten.
Man vergleiche einmal diese rasche Evolution von Weizen und Gerste mit dem Aufstieg von Mais, der wichtigsten Getreidepflanze der Neuen Welt. Der vermutete Maisvorfahr, eine Wildpflanze namens Teosinte, unterscheidet sich in Samen und Blütenbau so sehr von Mais, daß unter Botanikern lange umstritten war, ob Mais überhaupt von Teosinte abstammt. Der Wert dieser Pflanze als Nahrungslieferant dürfte Jäger und Sammler kaum sonderlich beeindruckt haben: Sie war in freier Natur nicht halb so ertragreich wie Wildweizen, produzierte viel weniger Samenkörner als der spätere Mais, und ihre Samen waren in harten, ungenießbaren Schalen verborgen. Damit aus Teosinte ein nützliches Anbaugewächs werden konnte, mußte sich ihre Reproduktionsbiologie grundlegend ändern, ihre Samen mußten viel größer werden, und die harten Schalen um die Körner mußten verschwinden. Unter Archäologen wird immer noch lebhaft darüber debattiert, wie viele Jahrhunderte oder sogar Jahrtausende wohl vergingen, bis die anfangs winzigen Maiskolben auch nur daumengroß waren. Klar erscheint indessen, daß es weitere Jahrtausende dauerte, bis Maiskolben von heutiger Größe geerntet werden konnten. Diese Diskrepanz zwischen den natürlichen Vorzügen von Weizen und Gerste und den Schwierigkeiten bei der Domestikation von Teosinte waren womöglich von entscheidender Bedeutung für die unterschiedliche Entwicklung der Gesellschaften in Eurasien und der Neuen Welt.
Ein dritter Vorteil der Pflanzenwelt Vorderasiens besteht darin, daß sie einen hohen Prozentsatz zwittriger Selbstbestäuber aufweist, das heißt von Pflanzen, die sich normalerweise selbst befruchten, wobei gelegentliche Kreuzbefruchtungen möglich sind. Wie schon erwähnt, sind die meisten Wildpflanzen entweder regelmäßig kreuzbefruchtete Zwitter oder sie treten in getrennter männlicher und weiblicher Form auf und sind auf gegenseitige Bestäubung angewiesen. Diese Fakten der Reproduktionsbiologie stellten frühzeitliche Bauern vor große Probleme, da immer dann, wenn sie einen ertragreichen Mutanten entdeckt hatten, dessen Nachkommen die geerbten Vorzüge durch Kreuzung mit anderen Pflanzen wieder verloren. Nicht zuletzt war dies der Grund, warum die meisten Kulturpflanzen zu dem geringen Prozentsatz von Wildpflanzen gehören, die entweder selbstbestäubende Zwitter sind oder sich durch ungeschlechtliche Fortpflanzung vermehren (z. B. durch Bildung genetischer Doppelgänger aus den Wurzeln einer Mutterpflanze). Der hohe Anteil zwittriger Selbstbestäuber an der Pflanzenwelt Vorderasiens stellte deshalb einen Vorteil für frühe Bauern dar, bedeutete er doch, daß ein hoher Prozentsatz der Wildpflanzen eine für den Menschen günstige Reproduktionsbiologie besaß.
Ein weiterer Vorteil von Selbstbestäubern lag darin, daß es bei ihnen von Zeit zu Zeit zu Kreuzbefruchtungen kam, wobei neue Sorten entstanden, unter denen dann ausgewählt werden konnte. Die gelegentlichen Kreuzbefruchtungen beschränkten sich nicht auf Pflanzen der gleichen Art, sondern bezogen auch verwandte Arten mit ein. Eine solche Artenkreuzung unter den Selbstbestäubern Vorderasiens brachte zum Beispiel den Brotweizen hervor, eine der wertvollsten Anbaupflanzen der Gegenwart.
Die ersten acht bedeutenden Anbaupflanzen, die in Vorderasien domestiziert wurden, waren ausnahmslos Selbstbestäuber. Von den drei Getreidearten darunter – Einkornweizen, Emmerweizen und Gerste – besaßen die beiden Weizensorten den zusätzlichen Vorteil eines hohen Eiweißgehalts (8–14 Prozent). Demgegenüber hatten die bedeutendsten Getreidepflanzen Ostasiens und der Neuen Welt – Reis und Mais – den Nachteil eines geringeren Eiweißgehalts.
Damit sind einige der Vorteile genannt, die Vorderasiens Pflanzenwelt für die ersten Bauern bereithielt: Sie wies einen ungewöhnlich hohen Anteil von Wildpflanzen auf, die sich zur Domestikation eigneten. Die mediterrane Klimazone, in der Vorderasien liegt, erstreckt sich jedoch nach Westen über einen großen Teil Europas und den Nordwesten Afrikas. Überdies weisen vier andere Regionen der Welt ebenfalls Zonen mit mediterranem Klima auf: Kalifornien, Chile, Südwestaustralien und Südafrika (Abbildung 7.2). Diese anderen Regionen waren aber nicht nur keine frühen Stätten der Landwirtschaft, sondern zählten überhaupt nicht zu den Orten, an denen sie unabhängig entstand. Was unterschied jene mediterrane Klimazone im Westen Eurasiens von allen anderen?
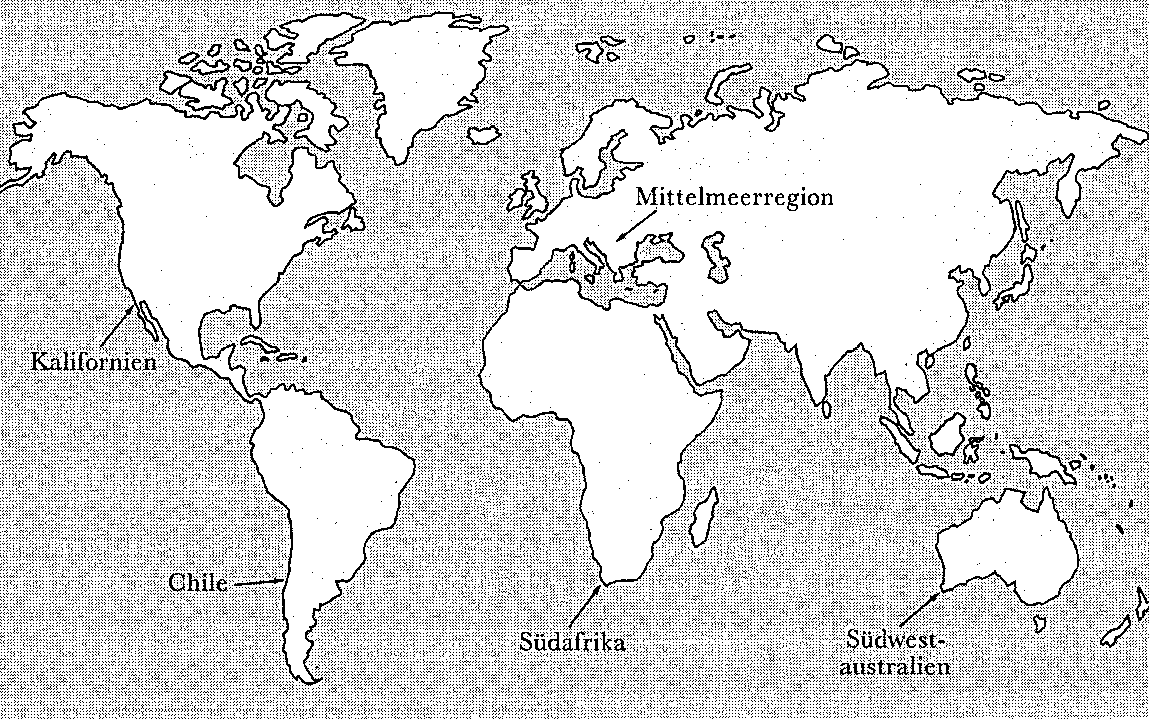
Abbildung 7.2 Die mediterranen Klimazonen der Welt
Wie sich herausstellt, hatte sie, und insbesondere ihr vorderasiatischer Teil, mindestens fünf Vorteile gegenüber anderen mediterranen Klimazonen. Erstens besitzt das westliche Eurasien bei weitem die größte Zone mit mediterranem Klima und weist deshalb eine viel größere pflanzliche und tierische Artenvielfalt auf als die vergleichsweise winzigen mediterranen Klimazonen Südwestaustraliens und Chiles. Zweitens sind die Klimaverhältnisse in Vorderasien von allen mediterranen Klimazonen am extremsten, mit starken Schwankungen zwischen den Jahreszeiten, aber auch von Jahr zu Jahr. Diese Schwankungen begünstigten die Evolution einjähriger Pflanzen, deren Anteil in diesem Raum besonders hoch ist. Die Kombination dieser ersten beiden Faktoren – große Artenvielfalt plus hohem Anteil einjähriger Pflanzen – führte dazu, daß die mediterrane Klimazone im westlichen Eurasien von allen Regionen der Welt mit Abstand die größte Vielfalt einjähriger Pflanzen aufweist.
Die Bedeutung dieser reichen botanischen Ausstattung für den Menschen wird durch die Untersuchungen des Geographen Mark Blumler über die Verbreitung von Wildgräsern veranschaulicht. Unter den Tausenden von Wildgräsern, die auf der Welt vorkommen, ermittelte Blumler die »Crème de la crème«, sprich die 56 Arten mit den größten Samen (mindestens zehnmal schwerer als der Durchschnitt aller Gräser), und faßte sie übersichtlich zusammen (siehe Tabelle 7.3). Wie sich zeigte, sind fast alle in mediterranen Klimazonen oder anderen Regionen mit periodischen Trockenzeiten heimisch. Außerdem ergab sich eine überwältigende Häufung im Bereich des Fruchtbaren Halbmonds und in anderen Teilen der mediterranen Klimazone des westlichen Eurasien, so daß angehende Bauern dort eine riesige Auswahl unter etwa 32 der 56 großsamigsten Wildgräser vorfanden! Zu erwähnen sind insbesondere Gerste und Emmerweizen, die beiden ältesten bedeutenden Kulturpflanzen Vorderasiens, die unter den 56 führenden Gräsern die Plätze 3 und 15 einnehmen. Demgegenüber fanden sich in der mediterranen Klimazone Chiles nur zwei dieser Arten, in Kalifornien und im südlichen Afrika nur jeweils eine und in Südwestaustralien überhaupt keine. Diese Tatsache allein ist schon von großer Tragweite für den Lauf der Geschichte.

Mark Blumler führt in seiner Doktorarbeit mit dem Titel »Seed Weight and Environment in Mediterraneantype Grasslands in California and Israel« (University of California, Berkeley, 1992) in Tabelle 12.1 die 56 Wildgräser mit den schwersten Samenkörnern (Bambus nicht mitgerechnet) auf, für die Daten vorlagen. Das Gewicht der Körner reichte bei den untersuchten Arten von 10 mg bis über 40 mg – das entspricht etwa dem Zehnfachen des mittleren Gewichts der Samenkörner aller Grasarten der Welt. Jene 56 Arten stellen weniger als ein Prozent aller Grasarten dar. Die obige Tabelle zeigt, daß sich diese »Supergräser« stark auf die mediterrane Zone des westlichen Eurasien konzentrieren.
Tabelle 7.1 Verbreitung großsamiger Gräser auf der Welt
Ein dritter Vorteil der mediterranen Klimazone in Vorderasien besteht in ihrer vielfältigen Topographie und den vielen verschiedenen Höhenlagen in geringer Entfernung voneinander. Das Spektrum reicht vom Toten Meer, dem niedrigsten Punkt der Erde, bis zu den über 5000 Meter hohen Berggipfeln unweit Teherans, mit der Folge, daß innerhalb kurzer Distanzen sehr unterschiedliche Umweltbedingungen herrschen, was wiederum zu einer großen Vielfalt von Wildpflanzen, den potentiellen Vorfahren unserer Kulturpflanzen, beitrug. Nicht weit von jenen steilen Berghängen liegen sanfte Flachlandzonen mit Flüssen, Überschwemmungsebenen und Wüsten, die durch Bewässerung fruchtbar gemacht werden konnten. Im Gegensatz hierzu herrscht in den mediterranen Klimazonen vor allem Südwestaustraliens, aber auch Südafrikas und Westeuropas weniger Abwechslungsreichtum in puncto Höhenlagen, Lebensräume und topographische Gegebenheiten.
Eine Folge der vielfältigen Höhenlagen im Bereich des Fruchtbaren Halbmonds waren gestaffelte Erntezeiten: Die Samen von Pflanzen, die in höheren Lagen wuchsen, wurden etwas später reif als die in tieferen Lagen. Sammler konnten deshalb bequem bergan ziehen und in jeder Höhenlage die gerade reif gewordenen Getreidekörner ernten, statt sich einem Meer von Gräsern mit gleichzeitig reifenden Körnern gegenüberzusehen, die gar nicht auf einmal zu bewältigen waren. Der Übergang zum Getreideanbau war dann gar nicht mehr so ein großer Schritt. Die ersten Ackerbauern brauchten bloß die Samen von Wildgetreide zu ernten, das an Berghängen wuchs, wo es auf unberechenbare Regenfälle angewiesen war, und diese dann in den feuchten Tälern auszusäen, damit sie dort, bei verringerter Abhängigkeit von Regenfällen, unter Aufsicht gedeihen konnten.
Mit der biologischen Vielfalt Vorderasiens auf engem Raum hängt auch ein vierter Vorteil zusammen: der Reichtum dieser Region nicht nur an Vorfahren wertvoller Kulturpflanzen, sondern auch an Vorfahren domestizierter Säugetiere. Wie wir sehen werden, kamen in den mediterranen Klimazonen Kaliforniens, Chiles, Südwestaustraliens und Südafrikas nur wenige oder gar keine Säugetierarten vor, die zur Domestikation geeignet waren. Demgegenüber wurden in Vorderasien vier Arten – Ziege, Schaf, Schwein und Rind – schon sehr früh domestiziert, vielleicht früher als irgendein anderes Tier an irgendeinem Ort der Welt (mit Ausnahme des Hundes). Diese vier Arten zählen noch heute zu den fünf wichtigsten Haustierarten (Kapitel 8). Ihre hauptsächlichen Verbreitungsgebiete lagen jedoch in verschiedenen Bereichen des Fruchtbaren Halbmonds, was dazu führte, daß sie auch an verschiedenen Orten domestiziert wurden: das Schaf wahrscheinlich im inneren Teil, die Ziege entweder in höhergelegenen Gebieten des östlichen Teils (iranisches Sagros-Gebirge) oder im Südwesten (Levante), das Schwein im nördlichen inneren Teil und das Rind im westlichen Teil einschließlich Anatoliens. Doch obwohl die Verbreitungsgebiete dieser vier ursprünglichen Vorfahren unserer Haustiere nicht identisch waren, lagen sie doch dicht genug beieinander, um nach erfolgter Domestikation einen Austausch zu ermöglichen, so daß am Ende alle vier Arten in der ganzen Region anzutreffen waren.
Der Aufstieg der Landwirtschaft in Vorderasien wurde durch die Domestikation von acht Anbaugewächsen ausgelöst, die wir als »Gründerpflanzen« bezeichnen (da sie die Landwirtschaft in der Region und vielleicht auf der ganzen Welt begründeten). Zu diesen acht Pflanzen zählten drei Getreidearten (Emmerweizen, Einkornweizen, Gerste), vier Hülsenfrüchte (Linse, Erbse, Kichererbse, Linsenwicke) und eine Faserpflanze (Flachs). Nur zwei der acht Gründerpflanzen, Flachs und Gerste, kommen in größerem Umfang auch außerhalb des Fruchtbaren Halbmonds und Anatoliens vor. Zwei weitere hatten ein sehr kleines natürliches Verbreitungsgebiet: Kichererbsen wuchsen nur im Südosten der Türkei und Emmerweizen nur im eigentlichen Gebiet des Fruchtbaren Halbmonds. Die entstehende Landwirtschaft konnte sich also auf die Domestikation heimischer Wildpflanzen stützen und war nicht auf den »Import« von Kulturpflanzen angewiesen. Außerdem konnten zwei der acht Gründerpflanzen nirgendwo anders domestiziert werden, da sie außer in Vorderasien nirgendwo auf der Welt heimisch waren.
Die Existenz geeigneter Wildtiere und -pflanzen versetzte die frühen Bewohner Vorderasiens in die Lage, innerhalb kurzer Zeit ein vielversprechendes und ausgewogenes Bio-Paket zu schnüren, das eine intensive Landwirtschaft ermöglichte. Es bestand aus drei Getreidearten als Hauptlieferanten von Kohlehydraten, vier Arten von Hülsenfrüchten mit 20–25 Prozent Eiweißgehalt und vier Haustierarten als Hauptlieferanten von Eiweiß, ergänzt durch den hohen Eiweißgehalt des Weizens, und Flachs als Faser- und Öllieferant (Leinsamen, wie die Flachssaat genannt wird, bestehen zu 40 Prozent aus Öl). Einige Tausend Jahre später wurden Tiere auch zur Gewinnung von Milch und Wolle, zum Pflügen von Feldern und als Transportmittel genutzt. Damit deckten die Anbaupflanzen und Tiere der ersten bäuerlichen Kulturen Vorderasiens schließlich die gesamten wirtschaftlichen Grundbedürfnisse des Menschen nach Kohlehydraten, Eiweiß, Fett, Kleidung, Zugkraft und Fortbewegungsmitteln.
Ein letzter Vorteil der frühen Landwirtschaft im Bereich des Fruchtbaren Halbmonds bestand darin, daß die Jagd- und Sammelwirtschaft dort möglicherweise weniger konkurrenzfähig war als in manchen anderen Regionen, so auch des westlichen Mittelmeerraums. Vorderasien hat nur wenige große Flüsse, und auch die Küste ist verhältnismäßig kurz, so daß die Möglichkeit, den Speiseplan mit Fischen und Schalentieren anzureichern, eher gering war. Eine wichtige Säugetierart, die wegen ihres Fleischs gejagt wurde, die Gazelle, kam ursprünglich in riesigen Herden vor, wurde aber parallel zum Anwachsen der menschlichen Bevölkerung stark dezimiert, bis nur noch ein kleiner Restbestand übrig war. So dauerte es nicht lange, bis die Landwirtschaft der Jagd- und Sammelwirtschaft überlegen war. Feste Dorfsiedlungen existierten zudem schon vor dem Aufkommen von Ackerbau und Viehzucht, so daß der Übergang zur Landwirtschaft für die Jäger und Sammler der Region naheliegender war als anderswo. In Vorderasien vollzog sich der Übergang innerhalb einer relativ kurzen Zeitspanne: Wurden noch um 9000 v. Chr. weder Pflanzen angebaut noch Haustiere gehalten, so waren einige Gesellschaften schon um 6000 v. Chr. vollständig auf domestizierte Pflanzen und Tiere angewiesen.
Ganz anders war die Situation in Mesoamerika: Dort gab es nur zwei domestizierbare Tiere (Truthahn und Hund), die obendrein sehr viel bescheidenere Fleischlieferanten waren als Rinder, Schafe, Ziegen und Schweine, und die wichtigste Getreideart, Mais, ließ sich, wie schon erwähnt, schwer domestizieren und womöglich auch nur sehr langsam durch Züchtung verändern. All dies führte dazu, daß die Domestikation in Mesoamerika wahrscheinlich erst um 3500 v. Chr. begann (der genaue Zeitpunkt ist noch umstritten); feste Dorfsiedlungen entstanden sogar erst um 1500 v. Chr.
Bei der gesamten bisherigen Erörterung der Vorzüge, die Vorderasien im Hinblick auf die frühe Entstehung der Landwirtschaft besaß, mußten wir kein einziges Mal irgendwelche vermeintlichen Vorzüge der Bewohner dieser Region ins Feld führen. Mir ist auch niemand bekannt, der ernsthaft behaupten würde, daß die Völker Vorderasiens bestimmte biologische Eigenschaften besaßen, die zum Erfolg ihres Bündels von Anbaupflanzen und Haustieren beigetragen haben könnten. Vielmehr haben wir gesehen, daß zahlreiche Einzelmerkmale des Klimas, der Umwelt, der Pflanzen- und Tierwelt Vorderasiens zusammen eine überzeugende Erklärung liefern.
Da die Kombinationen aus Kulturpflanzen und Haustieren, die sich in Neuguinea und im Osten der USA entwickelten, bei weitem nicht so effektiv waren wie in Vorderasien, könnte man vielleicht auf die Idee kommen, daß die dortigen Völker die Schuld daran trugen. Bevor wir uns näher mit den beiden Regionen beschäftigen, müssen wir uns mit zwei Fragen auseinandersetzen, die sich für alle Gebiete der Welt stellen, in denen die Landwirtschaft entweder gar nicht unabhängig entstand oder wo weniger hochkarätige Bündel aus Anbaupflanzen und Haustieren geschnürt wurden. Die erste Frage lautet, ob Jäger und Sammler und angehende Bauern alle örtlichen Wildpflanzen und ihre mögliche Verwendung wirklich gut kennen oder ob sie potentielle Vorfahren wertvoller Anbaupflanzen übersehen haben könnten. Falls ersteres zuträfe, würde die zweite Frage lauten, ob sie ihr Wissen auch tatsächlich zur Domestikation der nützlichsten Arten einsetzen oder ob kulturelle Faktoren sie daran hindern.
Was die erste Frage betrifft, widmet sich eine ganze Wissenschafts disziplin, die Ethnobiologie, der Erforschung der Wissensbestände verschiedener Völker über die Flora und Fauna ihrer jeweiligen Umgebung. Schwerpunkt dieser Studien sind und waren in erster Linie die wenigen überlebenden Jäger- und Sammlervölker der Erde sowie bäuerliche Gesellschaften, in denen Nahrung aus der Natur noch immer eine wichtige Rolle spielt. Die Untersuchungen kommen übereinstimmend zu dem Ergebnis, daß die Angehörigen dieser Völker geradezu wandelnde biologische Lexika sind, mit Namen (in der jeweiligen Sprache) für bis zu tausend oder mehr Pflanzen- und Tierarten und präzisem Wissen über die biologischen Merkmale jeder Art, ihre Verbreitung und möglichen Verwendungszwecke. Nimmt im Laufe der Geschichte die Abhängigkeit von domestizierten Pflanzen und Tieren ab, verlieren diese traditionellen Kenntnisse langsam an Wert und gehen verloren. Am Ende der Entwicklung steht der moderne Supermarktbesucher, der in der Natur ein Wildgras nicht von einer Hülsenfrucht unterscheiden kann.
Ich will ein typisches Beispiel geben. In den letzten 33 Jahren, in denen ich in Neuguinea immer wieder biologische Studien durchgeführt habe, war ich bei der Forschungsarbeit ständig in Begleitung von Neuguineern, für die Wildpflanzen und -tiere noch heute eine große Rolle spielen. Eines Tages, als ich mit meinen Gefährten vom Foré-Stamm im Dschungel ohne Nahrung festsaß, weil ein anderer Stamm uns den Rückweg zu unserem Lebensmitteldepot versperrte, kehrte ein Foré-Mann mit einem großen Rucksack voller Pilze, die er gesammelt hatte, zum Lager zurück und fing an, sie zu braten. Endlich wieder eine Mahlzeit! Doch dann kam mir ein beunruhigender Gedanke in den Sinn: Was, wenn die Pilze giftig waren? Geduldig erläuterte ich meinen Foré-Freunden, ich hätte gelesen, daß einige Pilzarten giftig seien, daß selbst einige amerikanische Pilzkenner gestorben seien, weil es so schwierig sei, Speise- von Giftpilzen zu unterscheiden, und daß es sich nicht lohne, das Risiko einzugehen, auch wenn wir alle noch so hungrig seien. An diesem Punkt wurden meine Gefährten ärgerlich und bedeuteten mir, ich solle ihnen jetzt einmal zuhören, sie würden mir gern einige Dinge klarmachen. Wie konnte ich sie, nachdem ich sie jahrelang über die Namen Hunderter von Bäumen und Vögeln ausgefragt hatte, jetzt so beleidigen, indem ich unterstellte, sie wüßten nicht die Unterschiede zwischen verschiedenen Pilzarten? Nur Amerikaner könnten so dumm sein, giftige mit eßbaren Pilzen zu verwechseln. Dann hielten sie mir noch Vorträge über 29 Arten von Speisepilzen, nannten für jede den Namen in der Foré-Sprache und erläuterten, wo man im Dschungel am besten nach ihnen suchen sollte. Der Pilz, der die Diskussion ausgelöst hatte, hieß Tánti, wuchs auf Bäumen und war besonders schmackhaft und absolut ungiftig.
Jedesmal, wenn ich Neuguineer auf Reisen in andere Teile ihrer Insel mitnehme, unterhalten sie sich häufig mit anderen Neuguineern, die uns begegnen, über die örtliche Natur und pflücken potentiell nützliche Pflanzen, um sie mit in ihre Heimatdörfer zu nehmen und dort versuchsweise anzupflanzen. Meine Erfahrungen mit Neuguineern decken sich mit denen von Ethnobiologen, die Völker mit traditioneller Lebensweise in anderen Regionen studiert haben. All diese Völker betreiben jedoch wenigstens in gewissem Umfang schon Landwirtschaft beziehungsweise sind die teilweise akkulturierten letzten Vertreter der alten Jäger- und Sammlerkulturen der Welt. Vor dem Aufstieg der Landwirtschaft, als alle Erdbewohner noch ausschließlich von dem lebten, was ihnen die Natur bot, war das Wissen über wilde Arten vermutlich noch viel umfassender. Die ersten Bauern waren Erben dieses Wissensschatzes, den im Laufe von Jahrzehntausenden Menschen, die in enger Gemeinschaft mit der Natur lebten, angehäuft hatten. Ich halte es deshalb für ausgesprochen unwahrscheinlich, daß potentiell nützliche Arten ihrer Aufmerksamkeit entgangen sind.
Die zweite, mit der ersten eng verknüpfte Frage lautet, ob vorgeschichtliche Sammler und Ackerbauern ihr ethnobiologisches Wissen bei der Auswahl und späteren Kultivierung geeigneter Wildpflanzen praktisch umsetzten. Eine Möglichkeit, dieser Frage nachzugehen, bietet die archäologische Fundstätte Tell Abu Hureyra am Rande des Euphrattals in Syrien. Zwischen 10000 und 9000 v. Chr. lebte die dortige Bevölkerung, so wird vermutet, schon ganzjährig in festen Siedlungen, betrieb jedoch noch Jagd- und Sammelwirtschaft. Der Übergang zur Landwirtschaft erfolgte erst ein Jahrtausend später. Die Archäologen Gordon Hillman, Susan Colledge und David Harris fanden an der Ausgrabungsstätte große Mengen verkohlter Pflanzenreste, bei denen es sich wahrscheinlich um weggeworfene Reste von Wildpflanzen handelte, die von den Bewohnern der Fundstätte gesammelt und dorthin gebracht worden waren. Die Wissenschaftler analysierten über 700 Proben, von denen jede im Durchschnitt mehr als 500 identifizierbare Samen von über 70 Pflanzenarten enthielt. Wie sich herausstellte, sammelten die Dorfbewohner eine ungeheure Vielzahl von Pflanzen (157 Arten!), die anhand ihrer verkohlten Samen bestimmt werden konnten, ganz zu schweigen von weiteren Pflanzen, deren Bestimmung heute nicht mehr möglich ist.
Sammelten jene naiven Dörfler wohl jede Art von Samenpflanze, deren sie habhaft werden konnten, trugen sie heim, vergifteten sich an den meisten Arten und ernährten sich von den wenigen übrigen? Sicher nicht. Während die Zahl von 157 Arten den Eindruck erwecken mag, als sei wahllos alles gesammelt worden, was nur zu finden war, fehlten unter den verkohlten Pflanzenresten doch viele Arten, die in der Umgebung ebenfalls wuchsen. Die 157 in den Proben enthaltenen Pflanzen fallen in drei Kategorien. Viele haben ungiftige, ohne weiteres eßbare Samen. Bei anderen, zum Beispiel einigen Hülsenfrüchten und Senfgewächsen, sind die Samen zwar giftig, doch das Gift läßt sich leicht entfernen, so daß die Samen doch noch verspeist werden können. Einige wenige Samen gehören zu Arten, die traditionell zum Färben oder als Heilmittel verwendet wurden. Die zahlreichen Wildpflanzen, die unter den 157 Arten nicht gefunden wurden, sind genau jene, die für den Menschen nutzlos oder schädlich waren, wie etwa alle besonders giftigen Kräuter, die in der Umgebung der Fundstätte anzutreffen waren.
Die Jäger und Sammler von Tell Abu Hureyra verschwendeten ihre Zeit also nicht und brachten sich auch nicht in Gefahr, indem sie wahllos alle möglichen Wildpflanzen heimtrugen. Sie waren mit der örtlichen Pflanzenwelt offenbar ebensogut vertraut wie moderne Neuguineer und setzten ihr Wissen ein, um nur die nützlichsten Samenpflanzen zu ernten. Eben diese bildeten aber vermutlich die Grundlage für die unbewußten ersten Schritte in Richtung Domestikation.
Schauplatz meines zweiten Beispiels dafür, wie vorgeschichtliche Völker ihr biologisches Wissen zum eigenen Vorteil nutzten, ist das Jordantal im 9. Jahrtausend v. Chr., als dort mit der Kultivierung von Anbaupflanzen begonnen wurde. Die ersten domestizierten Getreidearten des Flußtals waren Gerste und Emmerweizen, die noch heute zu den ertragreichsten Anbaupflanzen der Welt zählen. Doch wie in Tell Abu Hureyra muß es auch hier Hunderte anderer samentragender Wildpflanzenarten in der Umgebung gegeben haben, von denen 100 oder mehr eßbar gewesen und vor Beginn der Pflanzendomestikation regelmäßig von Sammlern geerntet worden sein dürften. Was war das Besondere an Gerste und Emmerweizen, das die Wahl ausgerechnet auf sie fallen ließ? Waren jene ersten Bauern des Jordantals botanische Stümper, die nicht wußten, was sie taten? Oder waren Gerste und Emmer tatsächlich die besten heimischen Wildgetreidearten der Region?
Zwei israelische Wissenschaftler, Ofer Bar-Yosef und Mordechai Kislev, bemühten sich um eine Antwort auf diese Frage, indem sie Wildgräser untersuchten, die im Jordantal noch heute vorkommen. Dabei ließen sie Arten mit kleinen oder ungenießbaren Samen außer acht und wählten nur die 23 wohlschmeckendsten, großsamigsten Wildgräser aus. Es überrascht nicht, daß sich darunter auch Gerste und Emmerweizen befanden.
Nun waren die 21 anderen Gräser auf der Liste allerdings nicht alle gleich nützlich. Von den 23 Arten erwiesen sich Gerste und Emmerweizen in vielerlei Hinsicht als die geeignetsten Kandidaten. Emmerweizen hat die größten Samenkörner, Gerste die zweitgrößten. In der Natur ist Gerste im Jordantal eines der vier Gräser mit der stärksten Verbreitung, Emmerweizen liegt im Mittelfeld. Zu den weiteren Vorteilen der Gerste zählen genetische und morphologische Eigenschaften, durch die nützliche Veränderungen in den natürlichen Samenverbreitungsmitteln und im Keimverzug (siehe letztes Kapitel) binnen relativ kurzer Zeit geschehen konnten. Emmerweizen hat dafür andere Vorzüge zu bieten: Er läßt sich besser ernten als Gerste und unterscheidet sich von den meisten anderen Getreidearten dadurch, daß die Körner fest von Spelzen umschlossen sind. Die Nachteile der 21 anderen Arten bestehen in kleineren Samen, geringerer natürlicher Verbreitung und zum Teil auch darin, daß es sich um perennierende statt um einjährige Pflanzen handelt, was Veränderungen im Zuge ihrer Domestikation stark verlangsamt hätte.
Die ersten Ackerbauern des Jordantals entschieden sich mit anderen Worten für die zwei besten der 23 geeignetsten Wildgräser, die in ihrer Umgebung wuchsen. Die evolutionären Veränderungen der Samenverbreitungsmittel und des Keimverzugs (nach Beginn der Kultivierung) waren natürlich unvorhergesehene Folgen des Handelns jener ersten Bauern. Die ursprüngliche Selektion von Gerste und Emmerweizen erfolgte aber bewußt und beruhte auf den leicht erkennbaren Merkmalen Samengröße, Genießbarkeit und natürliche Verbreitung. Das Beispiel des Jordantals verdeutlicht ebenso wie das von Tell Abu Hureyra, daß die ersten Bauern ihre ausführlichen Kenntnisse über heimische Arten zum eigenen Vorteil einzusetzen verstanden. Bei dem umfangreichen Wissen über die Pflanzenwelt ihrer Umgebung, wie sie heute höchstens noch eine kleine Zahl studierter Botaniker besitzt, konnte es ihnen kaum passieren, daß sie eine nützliche Wildpflanzenart übersahen und zu kultivieren versäumten, die ähnlich gut zur Domestikation geeignet war wie Gerste und Emmerweizen.
Wir wollen nun untersuchen, wie sich Bauern in zwei Regionen der Welt (Neuguinea, Osten der USA) mit unabhängig entstandener, aber anscheinend weniger effektiver Nahrungsproduktion (verglichen mit Vorderasien) verhielten, als ertragreichere Kulturpflanzen aus anderen Regionen eintrafen. Falls diese Pflanzen aus kulturellen oder sonstigen Gründen nicht übernommen wurden, müßten wir weiter nagende Zweifel hegen. Ungeachtet alles bisher Gesagten bliebe der Verdacht, in der lokalen Wildflora könnte sich vielleicht doch ein Vorfahr einer potentiell wertvollen Anbaupflanze verbergen, der von den örtlichen Bauern aufgrund kultureller Faktoren nicht beachtet wurde. Am Beispiel der beiden Regionen wird uns außerdem eine entscheidende Tatsache der Geschichte vor Augen geführt: daß nämlich Kultur pflanzen aus verschiedenen Teilen der Erde unterschiedlich ertragreich waren.
Neuguinea, nach Grönland die zweitgrößte Insel der Welt, liegt nur etwas nördlich von Australien in Äquatornähe. Wegen seines tropischen Klimas und der äußerst vielfältigen Lebensräume und Topographie besitzt Neuguinea eine reichhaltige Flora und Fauna (allerdings keine so reichhaltige wie vergleichbare festländische Tropenregionen). Die Besiedlung Neuguineas durch den Menschen reicht mindestens 40 000 Jahre zurück – viel länger als die Nord- und Südamerikas und etwas länger als die Besiedlung Westeuropas durch anatomisch weiterentwickelte Menschen. Insofern hatten die Neuguineer reichlich Gelegenheit, mit der örtlichen Pflanzen- und Tierwelt vertraut zu werden. Waren sie aber auch motiviert, ihr Wissen einzusetzen, um eine landwirtschaftliche Nahrungsproduktion aufzu bauen?
Ich erwähnte bereits, daß wir es beim Übergang zur Landwirtschaft mit einer Konkurrenzsituation zwischen dem Jagen und Sammeln auf der einen und Ackerbau und/oder Viehzucht auf der anderen Seite zu tun haben. In Neuguinea ist die Jagd- und Sammelwirtschaft nicht in dem Maße lohnend, daß der Anreiz zur Nahrungsproduktion dadurch geschmälert würde. Die heutigen Jäger und Sammler der Insel leiden besonders unter der ausgesprochenen Wildarmut: Unter den landbewohnenden Tierarten, die in Neuguinea heimisch sind, ist keins größer als der Kasuar, ein 100 Pfund schwerer flugunfähiger Vogel, und als ein 50 Pfund schweres Känguruh. Im neuguineischen Tiefland spielen in Küstennähe Fische und Schalentiere eine wichtige Rolle als Nahrungslieferanten, während viele Bewohner des Landesinneren noch als Jäger und Sammler leben und sich vorwiegend von wildwachsenden Sagopalmen ernähren. Von den Hochlandvölkern Neuguineas hat jedoch keins die Jagd- und Sammelwirtschaft bis in die Gegenwart beibehalten; sie leben durchweg in bäuerlichen Gesellschaften, die mit Nahrung aus der Natur lediglich ihren Speiseplan bereichern. Begeben sich Hochlandbewohner im Dschungel auf die Jagd, so nehmen sie als Proviant Gemüse mit, das aus ihren Gärten stammt. Falls sie das Pech haben, daß ihnen die Vorräte ausgehen, müssen sie trotz ihrer detaillierten Kenntnis der heimischen Wildpflanzen und -tiere im Wald verhungern. Da die Jagd- und Sammelwirtschaft mit anderen Worten in großen Teilen des heutigen Neuguinea unpraktikabel ist, überrascht es nicht, daß alle neuguineischen Hochland- und die meisten Tieflandbewohner heutzutage seßhafte Bauern sind. Große, früher bewaldete Gebiete des Hochlands wurden von traditionellen neuguineischen Bauern in abgezäunte, entwässerte, intensiv genutzte Ackerflächen verwandelt, die eine große Zahl von Menschen ernähren.
Archäologische Funde lassen den Schluß zu, daß die Landwirtschaft in Neuguinea schon vor langer Zeit begann, etwa um 7000 v. Chr. In jener Zeit waren alle Landmassen in der Umgebung der Insel noch ausschließlich von Jägern und Sammlern besiedelt, so daß die frühe Landwirtschaft auf Neuguinea unabhängig von äußeren Einflüssen entstanden sein muß. Zwar wurden bisher keine eindeutigen Überreste von Anbaupflanzen aus vorgeschichtlicher Zeit entdeckt, doch dürften einige der gleichen Pflanzen darunter gewesen sein, die zur Zeit der europäischen Kolonisation angebaut wurden und von denen man inzwischen weiß, daß sie wilde neuguineische Vorfahren besitzen. Von herausragender Bedeutung ist Zuckerrohr, die führende Anbaupflanze der heutigen Welt, deren Jahresproduktion in Tonnen fast so hoch ist wie die der Pflanzen auf den Rängen 2 und 3 (Weizen und Mais) zusammen. Andere Kulturpflanzen von unumstritten neuguineischer Herkunft sind unter anderem einige Bananenarten mit der Sammelbezeichnung Australimusa, der Nußbaum Canarium indicum, Sumpftaro sowie verschiedene eßbare Gräser, Wurzeln und grüne Gemüse. Brotfruchtbaum, Jamswurzel und (gewöhnlicher) Taro wurden möglicherweise ebenfalls in Neuguinea domestiziert, doch läßt sich darüber nur spekulieren, da die wildwachsenden Vorfahren dieser Gewächse außer in Neuguinea auch in Teilen Südostasiens vorkamen. Zur Zeit fehlen noch Hinweise, die eindeutig darüber Auskunft geben, ob die Domestikation dieser Arten in Südostasien, wie bisher angenommen, oder auch eigenständig in Neuguinea (oder sogar nur dort) erfolgte.
Bei näherer Untersuchung stellt man allerdings fest, daß Neuguineas Flora und Fauna von drei schwerwiegenden Nachteilen geprägt ist. Erstens wurde auf der Insel – im Gegensatz zu Vorderasien, der Sahelzone und China – kein einziges Getreide domestiziert. Mit der starken Konzentration auf Knollen- und Baumfrüchte verkörpert Neuguinea den Extremfall eines Phänomens, das auch in anderen feuchten Tropenregionen (Amazonasbecken, tropisches Westafrika, Südostasien) anzutreffen ist, wo ebenfalls Wurzelfrüchte wichtige Anbaugewächse darstellten; daneben wurden in diesen Regionen aber mindestens zwei Getreidearten (asiatischer Reis und Tränengras, ein besonders großkörniges asiatisches Getreide) domestiziert. Der Grund dafür, daß in Neuguinea keine Landwirtschaft auf Getreidebasis entstand, lag vermutlich darin, daß kein entsprechendes Rohmaterial vorhanden war: Von den 56 großsamigsten Wildgräsern der Welt ist kein einziges auf der Insel heimisch.
Zweitens war unter den Tieren Neuguineas kein einziges größeres Säugetier, das sich zur Domestikation geeignet hätte. Die einzigen Haustiere, die heute auf der Insel gehalten werden, Schwein, Huhn und Hund, trafen innerhalb der letzten Jahrtausende auf dem Weg über Indonesien vom südostasiatischen Festland her ein. Während die Bewohner des Tieflands ihren Eiweißbedarf durch Fischfang decken, leiden die bäuerlichen Hochlandbewohner wegen des niedrigen Eiweißgehalts ihrer Grundnahrungsmittel (Taro und Süßkartoffel) unter chronischem Eiweißmangel. So liegt der Eiweißgehalt von Taro bei nur 1 Prozent, also weit unter dem von Reis und noch weiter unter dem der Weizenkörner und Hülsenfrüchte des Fruchtbaren Halbmonds (8–14 beziehungsweise 20–25 Prozent).
Bei den Kindern im neuguineischen Hochland beobachtet man noch heute geschwollene Bäuche, ein typisches Anzeichen von Eiweißmangel, gepaart mit der Aufnahme großer Nahrungsmengen. Alte und junge Neuguineer verspeisen regelmäßig Mäuse, Spinnen, Frösche und anderes Kleingetier, das in anderen Regionen, wo große Haustiere oder Wild zur Verfügung stehen, als Nahrungsquelle verschmäht wird. Wahrscheinlich ist Eiweißmangel auch die tiefere Ursache des verbreiteten Kannibalismus in den traditionellen neuguineischen Hochlandkulturen.
Und schließlich lieferten die in Neuguinea in früherer Zeit angebauten Wurzelfrüchte nur begrenzte Kalorien- und Eiweißmengen, da sie in den großen Höhen, in denen viele Neuguineer heute leben, nicht gut gedeihen. Vor mehreren Jahrhunderten kam jedoch mit der Süßkartoffel, die ursprünglich aus Südamerika stammte und wahrscheinlich über die Philippinen, wohin sie die Spanier brachten, nach Neuguinea gelangte, eine neue Wurzelfrucht auf die Insel. Verglichen mit Taro und anderen älteren neuguineischen Anbaugewächsen gedeiht die Süßkartoffel auch in höheren Lagen, zeichnet sich durch schnelleres Wachstum aus und liefert höhere Hektarerträge, die noch dazu weniger Arbeitseinsatz erfordern. Die Folge des Imports der Süßkartoffel war eine Bevölkerungsexplosion im Hochland von Neuguinea. Das zeigt, daß die heimischen Kulturpflanzen, die vor dem Eintreffen der Süßkartoffel seit Jahrtausenden angebaut worden waren, sowohl der Bevölkerungsdichte als auch dem Siedlungsgebiet (Höhenlage) Grenzen gesetzt hatten.
Neuguinea bietet somit einen lehrreichen Gegensatz zu Vorderasien. Wie die Jäger und Sammler im Bereich des Fruchtbaren Halbmonds brachten auch die neuguineischen die Landwirtschaft eigenständig hervor. Ihr Anbausystem litt jedoch unter dem Fehlen domestizierbarer Getreidearten, Hülsenfrüchte und Tiere, dem daraus resultierenden Mangel an eiweißreicher Nahrung im Hochland sowie dem schlechteren Gedeihen heimischer Wurzelfrüchte in höheren Lagen. Dabei können sich die Kenntnisse der Neuguineer über die Wildpflanzen und -tiere ihrer Umwelt mit denen aller anderen Völker der Erde messen. Es darf getrost davon ausgegangen werden, daß sie alle Pflanzenarten, deren Domestikation lohnend gewesen wäre, entdeckt und getestet hätten. Auch sind die Neuguineer sehr wohl in der Lage, sinnvolle Ergänzungen ihres Sortiments an Anbaupflanzen zu erkennen, wie die eifrige Übernahme der Süßkartoffel beweist. Die gleiche Lehre vermitteln auch heutige Entwicklungen auf Neuguinea, wo Stämme mit leichterem Zugang zu importierten Anbaupflanzen und Haustieren (beziehungsweise höherer kulturell bedingter Übernahmebereitschaft) auf Kosten von Stämmen, denen es daran mangelt, expandieren. Der begrenzte Erfolg der Landwirtschaft auf Neuguinea hatte also nicht das geringste mit den Völkern Neuguineas zu tun, sondern rührte ausschließlich von Flora, Fauna und Umweltbedingungen her.
Mein zweites Beispiel dafür, wie eine unabhängig entstandene Landwirtschaft durch die Gegebenheiten der örtlichen Pflanzenwelt in ihrer Entwicklung beschränkt wurde, liefert der Osten der USA. Wie in Neuguinea wurden dort heimische Wildpflanzen eigenständig domestiziert. Allerdings ist die früheste Entwicklung im Osten der USA weitaus besser erforscht als in Neuguinea: Die Kulturpflanzen der ersten Ackerbauern konnten bestimmt werden, und auch Zeitpunkte und Reihenfolge der lokalen Domestikation sind bekannt. Lange vor der Ankunft von Anbaupflanzen aus anderen Gegenden siedelten sich nordamerikanische Indianer in Flußtälern an und betrieben dort auf der Grundlage lokaler Anbaugewächse eine intensive Landwirtschaft. Sie waren mithin in der Lage, die Vorzüge der vielversprechendsten Wildpflanzen zu nutzen. Welche Pflanzen kultivierten sie am Ende, und wie schnitt ihr »Gründerpaket« im Vergleich zu dem des Fruchtbaren Halbmonds ab?
Vier »Gründerpflanzen« wurden im Zeitraum zwischen 2500 und 1500 v. Chr. im Osten der USA domestiziert, volle 6000 Jahre nach der Domestikation von Weizen und Gerste in Vorderasien. Eine örtliche Kürbisart lieferte neben kleinen Behältern eßbare Kerne. Die drei übrigen Gründerpflanzen wurden nur wegen der eßbaren Kerne angebaut (Sonnenblume, ein Gänseblümchen-Verwandter namens Sumpfholunder und Gänsefuß, ein entfernter Verwandter des Spinats).
Drei Nahrungspflanzen und ein Gewächs zur Herstellung von Behältern ergaben jedoch noch keine ausreichende Grundlage für den Einstieg in die Landwirtschaft. Zwei Jahrtausende lang fristeten diese Gründerpflanzen deshalb ein relativ unbedeutendes Dasein als Ergänzung des Speiseplans der Indianerstämme im Osten der USA, die sich weiterhin in erster Linie von den Gaben der Natur ernährten (insbesondere von Wild und Wasservögeln, Fisch, Schalentieren und Nüssen). Erst ab 500–200 v. Chr., als noch drei weitere Anbaupflanzen (Knöterich, Maygrass und Little Barley, eine Gerstensorte) hinzugekommen waren, wuchs die Bedeutung der Landwirtschaft.
Ein moderner Ernährungswissenschaftler wäre von den sieben Anbaupflanzen des amerikanischen Ostens hellauf begeistert. Alle besaßen einen hohen Eiweißgehalt (17–32 Prozent gegenüber 8–14 Prozent bei Weizen, 9 Prozent bei Mais und noch weniger bei Gerste und weißem Reis). Zwei von ihnen (Sonnenblume und Sumpfgras) waren überdies sehr ölhaltig (45–47 Prozent). Vor allem Sumpfgras wäre mit 32 Prozent Eiweiß- und 45 Prozent Ölgehalt den Idealvorstellungen eines Ernährungswissenschaftlers sehr nahe gekommen. Wie kommt es dann, daß diese Idealkost von unserem Speiseplan so gänzlich verschwunden ist?
Bei den meisten der genannten Gewächse standen dem Vorteil eines hohen Nährwerts schwere Nachteile gegenüber. Gänsefuß, Knöterich, Little Barley und Maygrass besaßen winzige Samen, deren Volumen nur einem Zehntel dessen von Weizen- und Gerstenkörnern entsprach. Noch schwerer wogen die Nachteile beim Sumpfgras, einem windbestäubten Verwandten des Ambrosienkrauts, das als Heuschnupfenerreger verschrien ist. Wie Ambrosienkraut können auch Sumpfgraspollen Heuschnupfen auslösen, wenn die Pflanze in dichten Beständen wächst. Wen das noch nicht davon abbringt, Sumpfgras anzubauen, dem sei noch gesagt, daß dieses Gewächs einen starken Geruch verströmt, der nicht jedem zusagt, und daß Berührungen zu Hautreizungen führen können.
Kurz nach Beginn unserer Zeitrechnung gelangten Kulturpflanzen aus Mexiko über Handelsrouten in den Osten der heutigen USA. Mais traf um 200 n. Chr. ein, spielte aber noch etliche Jahrhunderte eine sehr untergeordnete Rolle. Um 900 n. Chr. tauchte endlich eine neue Maissorte auf, die an die kurzen Sommer in Nordamerika angepaßt war, und mit der Ankunft der Bohne um 1100 n. Chr. war das mexikanische Trio aus Mais, Bohne und Kürbis schließlich komplett. Die Landwirtschaft im Osten der USA erhielt dadurch kräftigen Auftrieb, und am Mississippi und seinen Nebenflüssen entstanden Reiche mit hoher Bevölkerungsdichte. In einigen Gebieten wurden die ursprünglich domestizierten Pflanzen parallel zu den viel ertragreicheren aus Mexiko weiter angebaut, in anderen jedoch vollständig von diesen verdrängt. Kein Europäer erblickte je Sumpfgras in indianischen Gärten, da diese Pflanze zur Zeit der europäischen Kolonisation Nord- und Südamerikas, die 1492 begann, nicht mehr kultiviert wurde. Unter allen erwähnten frühen Anbaupflanzen des amerikanischen Ostens konnten sich nur zwei (Sonnenblume und Östlicher Kürbis) in der Konkurrenz mit domestizierten Pflanzen fremder Herkunft behaupten und werden auch jetzt noch angebaut. Verschiedene heutige Kürbisarten sind direkte Nachfahren jener schon vor Tausenden von Jahren domestizierten amerikanischen Kürbisse.
Der Osten der USA hält somit, wie Neuguinea, einige interessante Lehren bereit. Grundsätzlich hätte man dort die unabhängige Entstehung einer ertragreichen Landwirtschaft für wahrscheinlich halten können. Die Region besitzt fruchtbare Böden, es fällt genügend, aber nicht zuviel Niederschlag, und das Klima ist immerhin so günstig, daß den heutigen Farmern reiche Erträge beschert werden. Die Pflanzenwelt ist artenreich und besteht unter anderem aus wilden Nußbäumen wie Eiche und Hickory. Die indianischen Bewohner der Region begründeten die Landwirtschaft auf der Basis heimischer Pflanzen, die sie domestizierten, und wurden in Dörfern seßhaft; im Zeitraum zwischen 200 v. Chr. und 400 n. Chr. kam es sogar zu einer kulturellen Blüte (Hopewell-Kultur im heutigen Ohio). Somit bestand mehrere Jahrtausende lang Gelegenheit, die nützlichsten Wildpflanzen der Region ausfindig zu machen und gegebenenfalls als Anbaupflanzen zu nutzen.
Wir dürfen aber nicht vergessen, daß die Hopewell-Blütezeit erst rund 9000 Jahre nach Entstehung der ersten dörflichen Gemeinschaften in Vorderasien anbrach. Und selbst dann dauerte es noch bis etwa 900 n. Chr., bis das mexikanische Pflanzentrio ein starkes Bevölkerungswachstum auslöste, in dessen Folge am Mississippi eine kulturelle Blütezeit begann, in welcher die größten städtischen Siedlungen und die hochentwickeltsten Gesellschaften entstanden, die uns von Indianern nördlich Mexikos bekannt sind. Doch dieser Boom kam viel zu spät, um die Indianer Nordamerikas für die herannahende Katastrophe der europäischen Kolonisation zu wappnen. Eine Landwirtschaft, die allein auf den Pflanzen des amerikanischen Ostens basierte, hatte aus naheliegenden Gründen nicht ausgereicht, um den Wachstumsschub auszulösen. Die heimischen Wildgetreide der Region waren nicht annähernd so potent wie Weizen und Gerste. Die Indianer des amerikanischen Ostens domestizierten auch niemals heimische Hülsenfrüchte, Faserpflanzen, Obst oder Nußbäume. Zudem besaßen sie außer Hunden, deren Domestikation wahrscheinlich in einem anderen Teil Nord- oder Südamerikas erfolgte, kein einziges Haustier.
Sicher ist auch, daß die Indianer im Osten der USA unter den Wildpflanzen ihrer Umgebung keine bedeutenden potentiellen Anbaupflanzen übersahen. Selbst Agrarwissenschaftler des 20. Jahrhunderts konnten, gestützt auf die gesammelten Erkenntnisse der modernen Forschung, nur geringe Erfolge bei der Nutzbarmachung nordamerikanischer Wildpflanzen verbuchen. Es stimmt zwar, daß Pekannüsse und Blaubeeren domestiziert und einige eurasische Obstpflanzen (Äpfel, Pflaumen, Weinbeeren, Himbeeren, Brombeeren, Erdbeeren) durch Kreuzung mit ihren wilden nordamerikanischen Verwandten verbessert wurden. Diese wenigen Erfolge änderten jedoch unsere gegenwärtigen Ernährungsgewohnheiten bei weitem nicht so nachhaltig, wie die der Indianer im Osten der USA durch die Einführung von Mais aus Mexiko in der Zeit nach 900 n. Chr. verändert wurden.
Diejenigen, die am meisten über die domestizierten Pflanzen des amerikanischen Ostens wußten, nämlich die dortigen Indianer, fällten ihr Urteil, indem sie sich nach Ankunft des mexikanischen Pflanzentrios ganz oder teilweise von ihnen abwandten. Das beweist zugleich, daß kein kultureller Konservatismus die amerikanischen Indianer davon abhielt, den Wert einer Anbaupflanze, die ihnen zu Gesicht kam, zu erkennen und sie zu übernehmen. Ähnlich wie in Neuguinea war auch im Osten der heutigen USA der beschränkte Erfolg der eigenständigen Landwirtschaft nicht der indianischen Bevölkerung zuzuschreiben, sondern gänzlich der amerikanischen Fauna, Flora und Umwelt.
Wir haben bisher drei Regionen mit unabhängig entstandener Landwirtschaft erörtert. Von ihnen ist der Fruchtbare Halbmond der eine Extremfall, den anderen bilden Neuguinea und der Osten der USA. Wir haben gesehen, daß die Völker Vorderasiens mit Abstand als erste mit der Domestikation von Pflanzen begannen. Sie domestizierten weit mehr Pflanzen, viel ertragreichere oder nützlichere Arten und ein erheblich breiteres Spektrum von Anbaugewächsen. Zudem entwickelten sie intensivere Formen der Landwirtschaft, die höhere Bevölkerungsdichten ermöglichten. Das alles geschah in größerem Tempo als anderswo, was letzten Endes dazu führte, daß die Völker Vorderasiens mit höher entwickelter Technik, komplexeren Formen politischer Organisation und einer größeren Zahl epidemischer Krankheiten, mit denen sie andere Völker anstecken konnten, ins moderne Zeitalter eintraten.
Wir haben festgestellt, daß diese Unterschiede zwischen dem Bereich des Fruchtbaren Halbmonds, Neuguinea und dem Osten der USA eindeutig aus der vorhandenen Ausstattung mit Wildpflanzen und -tieren resultierten, die zur Domestikation zur Verfügung standen, nicht aber aus besonderen Merkmalen der dortigen Völker. Von außen eingeführte ertragreichere Kulturpflanzen (Süßkartoffel in Neuguinea, mexikanisches Pflanzentrio im Osten der USA) wurden prompt übernommen, mit der Folge einer Intensivierung der Landwirtschaft und eines steilen Bevölkerungsanstiegs. Entsprechend gehe ich davon aus, daß in Regionen der Erde, in denen die Landwirtschaft nicht unabhängig entstand – wie Kalifornien, Australien, die argentinische Pampa und Westeuropa – wahrscheinlich noch weniger domestizierbare Wildpflanzen und -tiere vorkamen als in Neuguinea und im Osten der USA, wo die Landwirtschaft wenigstens ein bescheidenes Niveau erreichte. Mark Blumlers bereits zitierte Analyse der weltweiten Verbreitung großsamiger Wildgräser sowie die Untersuchung der Verbreitung großer Säugetiere, die Gegenstand des nächsten Kapitels sein wird, ergeben übereinstimmend, daß in all jenen Regionen, in denen die Landwirtschaft gar nicht entstand oder nie über das Anfangsstadium hinauskam, ein Mangel an Wildvorfahren domestizierbarer Vieh- und Getreidearten herrschte.
Erinnern wir uns, daß in der Anfangsphase der Landwirtschaft auch die Konkurrenz zwischen Nahrungsproduktion auf der einen und Jagd- und Sammelwirtschaft auf der anderen Seite eine Rolle spielte. Man könnte deshalb fragen, ob nicht überall dort, wo die Landwirtschaft nur langsam oder gar nicht entstand, ein besonderer Ressourcenreichtum, der günstige Voraussetzungen für die Jagd- und Sammelwirtschaft schuf, die Ursache war, nicht aber eine besonders geringe Zahl domestizierbarer Arten. In Wirklichkeit war es so, daß die meisten dieser Regionen Jägern und Sammlern besonders wenig zu bieten hatten, da die meisten großen Säugetiere Australiens und Nord- und Südamerikas (anders als in Eurasien und Afrika) gegen Ende des Eiszeitalters ausgestorben waren. Die Landwirtschaft wäre somit in diesen Regionen auf noch weniger Konkurrenz gestoßen als in Vorderasien. Erstklassige Jagdreviere können demnach nicht dafür verantwortlich gemacht werden, wenn die Landwirtschaft nicht entstand oder in den Kinderschuhen verharrte.
Um Mißverständnissen vorzubeugen, sei an dieser Stelle noch einmal hervorgehoben, daß weder die Bereitschaft zur Übernahme besserer Anbaupflanzen und Haustiere noch die Beschränkungen aufgrund des lokalen Angebots an Wildpflanzen und -tieren überbetont werden sollten. Beides sind keine absoluten Gegebenheiten. Wir haben bereits zahlreiche Beispiele erörtert, bei denen ertragreichere Anbaupflanzen fremder Herkunft übernommen wurden. Daraus kann geschlossen werden, daß Völker in der Lage sind, nützliche Pflanzen zu erkennen, daß sie deshalb besser geeignete domestizierbare Pflanzen – sofern vorhanden – wahrscheinlich auch erkannt hätten und nicht durch kulturellen Konservatismus oder Tabus daran gehindert wurden. Einer wichtigen Ergänzung bedarf diese Aussage jedoch: All dies gilt nur bei langfristiger und großräumiger Betrachtung. Jeder Kenner der Geschichte menschlicher Kulturen kann nämlich zahllose Beispiele anführen, bei denen Anbaupflanzen, Haustiere und andere Neuerungen abgelehnt wurden, obwohl sie durchaus nützlich gewesen wären.
Natürlich hege ich nicht die naive Vorstellung, daß jede Gesellschaft sofort jede Neuerung übernimmt, die für sie von Vorteil wäre. Tatsache ist vielmehr, daß bei Betrachtung ganzer Kontinente und anderer geographischer Großräume mit Hunderten konkurrierender Gesellschaften einige dieser Gesellschaften eine aufgeschlossenere Haltung gegenüber Neuerungen einnehmen und andere eine ablehnendere. Die aufgeschlosseneren Gesellschaften, die sich für die Übernahme neuer Anbaupflanzen, Haustiere oder Techniken entscheiden, sind wahrscheinlich diejenigen, die sich besser ernähren, rascher vermehren und am Ende innovationsscheue Gesellschaften verdrängen oder gar vernichten. Wir haben es hier mit einem wichtigen Phänomen zu tun, dessen Erscheinungsformen weit über die Einführung neuer Kulturpflanzen hinausgehen und auf das wir in Kapitel 12 zurückkommen werden.
Überbetont werden sollten auch nicht die Beschränkungen, die sich aus dem vorhandenen Angebot an Wildpflanzen und -tieren für die Entstehung der Landwirtschaft ergeben. Ich sage ausdrücklich nicht, daß die Landwirtschaft in jenen Regionen, in denen sie bis in die jüngere Vergangenheit nicht eigenständig hervorgebracht wurde, nie von selbst entstanden wäre. Wenn heute davon gesprochen wird, daß die australischen Aborigines bei Anbruch der Neuzeit noch in der Steinzeit lebten, steht dahinter oft die Vorstellung, daß dieser Zustand ewig fortgedauert hätte.
Um den darin liegenden Trugschluß zu begreifen, stellen wir uns einmal vor, ein Besucher aus dem All würde der Erde im Jahr 3000 v. Chr. einen Besuch abstatten. Der Außerirdische fände im Osten der USA keinerlei Spuren der Landwirtschaft vor, weil diese in dem Gebiet erst um 2500 v. Chr. auf den Plan trat. Zöge der Besucher im Jahr 3000 v. Chr. den Schluß, negative Merkmale der Pflanzen- und Tierwelt im Osten der USA würden die Entstehung der Landwirtschaft dort für alle Zeit unmöglich machen, hätten ihn die Ereignisse im Jahrtausend darauf Lügen gestraft. Selbst ein Besucher Vorderasiens hätte zu dem Fehlurteil verleitet werden können, diese Region sei für die Landwirtschaft verloren, wenn seine Visite im Jahr 9500 v. Chr. und nicht erst 8500 v. Chr. stattgefunden hätte.
Meine These lautet demnach nicht, daß es in Kalifornien, Australien, Westeuropa und all den anderen Gebieten, in denen die Landwirtschaft nicht unabhängig entstand, keine domestizierbaren Arten gab und daß dort für immer nur Jäger und Sammler gelebt hätten, wenn keine fremden Völker oder an fremden Orten domestizierten Pflanzen und Tiere eingetroffen wären. Vielmehr stelle ich fest, daß zwischen den Regionen große Unterschiede in der Zahl der vorhandenen domestizierbaren Arten bestanden, daß sich die Regionen entsprechend auch im Zeitpunkt der Entstehung der Landwirtschaft unterschieden und daß in einigen fruchtbaren Regionen bis in die Neuzeit hinein keine Nahrungsproduktion unabhängig entstand.
Australien, der vermeintlich »rückständigste« Kontinent, liefert hierfür ein gutes Beispiel. Im mit Wasser reich gesegneten Südosten, dem für die Landwirtschaft am besten geeigneten Teil Australiens, schlugen örtliche Aborigines-Gesellschaften offenbar in den letzten Jahrtausenden einen Weg ein, der irgendwann einmal zur Nahrungsproduktion geführt hätte. So hatten sie bereits Winterdörfer errichtet und damit begonnen, ihre Umwelt systematisch zu bewirtschaften, indem sie Fischfallen, Netze und sogar lange Kanäle bauten. Hätten die Europäer Australien nicht 1788 kolonisiert und dieser Entwicklung ein Ende gesetzt, wären australische Aborigines möglicherweise binnen weniger Jahrtausende zu Nahrungsproduzenten geworden, mit Fischteichen und Feldern, auf denen domestizierte australische Jamswurzeln und kleinsamige Gräser wuchsen.
Vor diesem Hintergrund können wir nun auch die in der Überschrift dieses Kapitels gestellte Frage beantworten, ob nämlich der Grund dafür, daß die nordamerikanischen Indianer keine Äpfel domestizierten, bei ihnen oder bei den Äpfeln zu suchen sei.
Es soll nicht unterstellt werden, daß Äpfel in Nordamerika nie hätten domestiziert werden können. Bedenken Sie, daß Apfelbäume historisch zu den am schwersten domestizierbaren Obstbäumen zählten und wegen der komplizierten Veredelungsverfahren, die dazu erforderlich waren, auch in Eurasien erst sehr spät domestiziert wurden. Es gibt keine Anzeichen, die dafür sprechen, daß mit der Kultivierung von Äpfeln auf breiter Front selbst in Vorderasien und Europa früher begonnen wurde als zur Zeit des klassischen Hellenismus, sprich 8000 Jahre nach dem Aufkommen der Landwirtschaft in Eurasien. Hätten die amerikanischen Indianer im gleichen Tempo Veredelungstechniken erfunden beziehungsweise übernommen, wären auch sie am Ende zur Domestikation von Äpfeln gelangt – etwa im Jahr 5500 n. Chr., also rund 8000 Jahre nach dem Beginn der Pflanzendomestikation in Nordamerika um 2500 v. Chr.
Die »Schuld« daran, daß die nordamerikanischen Indianer vor dem Eintreffen der Europäer keine nordamerikanischen Äpfel domestizierten, ist also weder den Indianern noch den Äpfeln anzulasten. Was die biologischen Voraussetzungen der Apfeldomestikation betrifft, so unterschieden sich weder indianische Bauern noch nordamerikanische Äpfel von ihren eurasischen Pendants. Einige der Supermarktäpfel, die Lesern dieses Kapitels vielleicht besonders schmackhaft erscheinen, wurden sogar erst vor relativ kurzer Zeit durch Kreuzung eurasischer und wilder nordamerikanischer Apfelsorten gezüchtet. Der Grund für das Ausbleiben der Apfeldomestikation in Nordamerika lag vielmehr in der gesamten Pflanzen- und Tierwelt, die von den dortigen Bewohnern vorgefunden wurde. Ihr vergleichsweise geringes Domestikationspotential erklärt den späten Start der Landwirtschaft in Nordamerika.