CINCO
MONTAJE DE UNA EXPLICACIÓN
UNA ALIANZA MISTERIOSA
El estudio de pacientes con recientes dificultades de razonamiento y toma de decisiones, descrito en la primera parte, condujo a identificar un conjunto específico de sistemas cerebrales que en esas personas estaban dañados. También permitió identificar una colección aparentemente extraña de procesos neuropsicológicos que dependen de que esos sistemas estén íntegros. ¿Qué interconecta esos procesos? ¿Y qué los vincula con los sistemas neurales que describí en el capítulo anterior? Los párrafos siguientes ofrecen algunas respuestas provisorias.
Primero, adoptar una decisión —compleja y de resultado incierto— en el típico problema personal que se plantea en un entorno social, requiere conocimientos amplios y estrategias racionales para operar sobre ese conocimiento. Este incluye hechos acerca de objetos, personas y situaciones en el mundo externo. Pero las decisiones personales y sociales son inextricables de la supervivencia, y entonces el conocimiento también incluye hechos y mecanismos relativos a la regulación del organismo como un todo. Los procesos de razonamiento se desenvuelven alrededor de opciones para la acción, predicción de futuros resultados y planes para implementar diversas metas en variadas escalas de tiempo.
Segundo, los procesos de emoción y sentimiento son parte de la maquinaria neural para la regulación biológica, cuyo núcleo está constituido por controles homeostáticos, pulsiones e instintos.
Tercero, debido al diseño del cerebro, el amplio conocimiento básico requerido depende de numerosos sistemas, situados en regiones cerebrales relativamente distantes y no tanto en una sola zona. Gran parte de ese conocimiento se recuerda en forma de imágenes en muchas localizaciones cerebrales y no tanto en una sola. Aunque tenemos la ilusión de que todo se reúne en un sólo teatro anatómico, recientes indicios sugieren que no es así. Es probable que la relativa simultaneidad de actividad en distintas localizaciones reúna las partes separadas de la mente.
Cuarto, como el conocimiento sólo se puede recuperar de modo distribuido y parcelado a partir de localizaciones situadas en muchos sistemas paralelos, la operación de las estrategias del razonamiento requiere que miles de hechos se representen activamente en un amplio despliegue paralelo durante un lapso considerable (de varios segundos por lo menos). En otras palabras, las imágenes con las cuales razonamos (imágenes de objetos específicos, actos y esquemas relaciónales; de palabras que ayudan a traducir estos últimos en lenguaje) no sólo deben estar «en foco» —lo que logra la atención—, sino que permanecer «activas en la mente», lo que consigue la memoria operativa.
Sospecho que la misteriosa alianza de los procesos descubiertos al final del capítulo anterior se debe parcialmente a la naturaleza de los problemas que el organismo intenta resolver y en parte al diseño del cerebro. Las decisiones personales y sociales están cargadas de incertidumbre e impactan —directa o indirectamente— en nuestras posibilidades de supervivencia. Requieren, entonces, un amplio repertorio de conocimientos relativos al mundo externo y al que contiene el organismo. Sin embargo, como el cerebro guarda y recupera el conocimiento de manera espacialmente separada y no de modo integrado, también necesitan atención y memoria operativa para que el componente del conocimiento —recuperado como despliegue de imágenes— se pueda manipular en el tiempo.
Sospecho, además, que una conveniencia evolutiva hace que los sistemas neurales identificados se superpongan de manera tan evidente. Si una fundamental regulación biológica es imprescindible para guiar la conducta social y personal, parece posible entonces que en el proceso de selección natural haya prevalecido un diseño cerebral en que los subsistemas responsables de lo racional-decisorio permanezcan íntimamente entretejidos con los que se ocupan de la regulación biológica, visto que comparten el logro de la supervivencia.
Este anticipo de explicación general constituye una primera aproximación a las interrogantes que plantea el caso Phineas Gage. ¿Qué hay en el cerebro que permite que los humanos se comporten racionalmente? ¿Cómo funciona? Suelo negarme a resumir el esfuerzo por contestar estas preguntas en la expresión «neurobiología de la racionalidad», porque suena oficial y pretenciosa, pero eso es, en suma: los comienzos de una neurobiología de la racionalidad humana en el nivel de los sistemas cerebrales de gran escala.
En la segunda parte de este libro, quiero poner a prueba la validez de la explicación general que acabo de dar y deducir de ella una hipótesis comprobable. Sin embargo, debido a la amplitud del tema, restringiré la discusión a una selección de materias que me parecen indispensables para que estas ideas resulten comprensibles.
Este capítulo es un puente entre los hechos de la primera parte y las interpretaciones que daré más adelante. El recorrido —espero que no se considere una interrupción— tiene varios propósitos: revisar ciertas nociones a las cuales recurriré frecuentemente (por ejemplo, organismo, cuerpo, cerebro, conducta, mente, estado); discutir brevemente la base neural del conocimiento destacando su naturaleza parcelada y su dependencia de imágenes, y comentar el desarrollo neural. No seré exhaustivo (por ejemplo, una discusión del aprendizaje o del lenguaje, sería apropiada y útil, pero ninguno de los dos tópicos es indispensable para mi actual objetivo). No ofreceré estudios bibliográficos de ningún tema, y no justificaré cada opinión que exprese. Recordemos que esta es una conversación.
Los capítulos subsiguientes retornan al relato principal, y se ocuparán de las regulaciones biológicas, de su expresión en emociones y sentimientos, y de los mecanismos por medio de los cuales emociones y sentimientos se utilizan en la toma de decisiones.
Antes de proseguir, quiero repetir algo que dije en la introducción: el texto es una indagación abierta más que un catálogo de hechos aceptados. No estoy haciendo afirmaciones que impliquen certeza absoluta, sino considerando hipótesis y pruebas empíricas.
DE ORGANISMOS, CUERPOS Y CEREBROS
Preguntemos lo que preguntemos sobre quiénes somos o por qué somos como somos, es evidente que somos organismos vivientes y complejos, con un cuerpo y un sistema nervioso («cerebro», para abreviar). Siempre que digo cuerpo me refiero al organismo sin incluir el tejido neural (los componentes centrales y periféricos del sistema nervioso), aunque el cerebro también es, en sentido convencional, parte del cuerpo.
El organismo tiene una estructura, y miles de componentes: un esqueleto óseo con un gran número de partes, conectado por articulaciones y movido por músculos; diversos órganos combinados en sistemas; una membrana que envuelve y delimita su frontera externa, construida principalmente de piel. De vez en cuando llamaré a los órganos —vasos sanguíneos, órganos en el pecho, abdomen y cabeza, piel— «víscera» (singular «viscus»). Nuevamente, en la acepción habitual, el cerebro estaría incluido, pero aquí lo excluyo.
Cada parte del organismo está hecha de tejidos biológicos, que a su vez están hechos de células. Cada una es una agregación de numerosas moléculas ordenadas para conformar un esqueleto para la célula (citoesqueleto), de muchos órganos y sistemas (núcleos celulares y órganos diversos), y un límite global (membrana celular). Cuando observamos una célula en actividad, nos amedrentan su compleja estructura y funcionamiento, y resulta asombroso ver funcionar un sistema orgánico corporal.
ESTADOS DE ORGANISMOS
En la exposición que sigue hay numerosas referencias a «estados corporales» y «estados mentales». Los organismos vivientes cambian continuamente, asumen una sucesión de «estados», cada uno de los cuales se define por diversas pautas de actividad en cada uno de sus componentes. Podemos imaginar esto como la suma de actividades de un montón de gente y objetos que trabajan en una zona circunscrita. Imagínate en la terminal de un gran aeropuerto, mirando afuera y adentro; ves y oyes el murmullo constante de diferentes sistemas: personas que ingresan o abandonan los aviones, otras sentadas o de pie; gente que camina o deambula con aparente decisión; aviones en la losa, carreteando, despegando, aterrizando; mecánicos y funcionarios que hacen su trabajo. Imagina ahora que congelas la imagen de ese video imaginario, o que fotografías la escena completa. Verás entonces la reproducción de un estado, un fragmento instantáneo, artificial de vida, que indica lo que estaba sucediendo en los diferentes componentes de un vasto organismo en el lapso que definió la velocidad del obturador de la cámara fotográfica. (En realidad, las cosas son algo más complicadas: según la escala de análisis, los estados de los organismos pueden ser unidades discretas o mezclarse continuamente).
CUERPO Y CEREBRO INTERACTÚAN: EL ORGANISMO POR DENTRO
Cuerpo y cerebro están indisociablemente integrados por circuitos bioquímicos y neurales que se apuntan unos a otros. La interconexión se verifica mediante dos rutas principales. La primera que se nos viene a la mente está hecha de nervios periféricos sensoriales y motores, que transmiten señales de todas las partes del cuerpo al cerebro y viceversa. La otra ruta, más difícil de imaginar aunque sea evolutivamente mucho más antigua, es el torrente sanguíneo: transporta señales químicas como las hormonas, los moduladores y los neurotransmisores.
Hasta un simplificado resumen pone de manifiesto lo complejo de estas interrelaciones:
- Prácticamente todas las partes del cuerpo, cada músculo, articulación y órgano interno, puede enviar señales al cerebro mediante los nervios periféricos. Estas señales ingresan al cerebro en el nivel de la médula espinal o del tronco del encéfalo y finalmente son transportados al interior del cerebro —pasando por diferentes postas neurales—, a las capas corticales somatosensoriales del lóbulo parietal y de la ínsula.
- La actividad corporal produce sustancias químicas que pueden llegar al cerebro por el torrente sanguíneo e influir la actividad cerebral directamente o por la activación de localizaciones encefálicas específicas, como el órgano subfornical.
- En la dirección opuesta, el cerebro puede actuar sobre distintas partes del cuerpo a través de los nervios. Los agentes para esas acciones son el sistema nervioso autónomo (o visceral) y el sistema nervioso musculoesquelético (o voluntario). Las señales para el sistema nervioso autónomo surgen en las regiones evolutivamente más antiguas (amígdala, corteza cingular, hipotálamo y tronco encefálico), en tanto que las señales para el sistema musculoesquelético se originan en varias capas corticales motoras y núcleos subcorticales motores de diferentes edades evolutivas.
- El cerebro también actúa sobre el cuerpo fabricando —u ordenando fabricar— sustancias químicas liberadas en el torrente sanguíneo, entre ellas las hormonas, los transmisores y los moduladores. Diré más sobre ellas en el próximo capítulo.
No exagero cuando digo que cuerpo y cerebro conforman un organismo indisociable. En realidad, estoy simplificando en exceso. Consideremos, por ejemplo, que el cerebro no sólo recibe señales desde el cuerpo, sino que —en algunos sectores— recepta señales de partes de sí mismo, partes que a su vez reciben señales desde el cuerpo. El organismo —constituido por la asociación cuerpo-cerebro— interactúa con el entorno como un conjunto, no siendo dicha interacción resultado de la acción independiente del cuerpo o del cerebro. Pero eso no es todo: los organismos complejos, como el nuestro, no sólo interactúan o generan las meras respuestas espontáneas o reactivas externas que colectivamente se conocen como conducta; también producen respuestas internas, algunas de las cuales constituyen las imágenes —visuales, auditivas, somatosensoriales, por ejemplo— que postulo como base de la mente.
DE LA CONDUCTA Y LA MENTE
Numerosos organismos simples —incluso aquellos que constan de una sola célula y no tienen cerebro— realizan acciones espontáneas o reactivas ante estímulos del entorno; es decir, producen conducta. Algunas de esas acciones están contenidas en el organismo mismo y pueden quedar ocultas para el observador (por ejemplo, la contracción en un órgano interno), o ser externamente observables (un espasmo, la extensión de un miembro). Otras acciones (reptar, caminar, sujetar algo) se dirigen al entorno. Pero en algunos organismos simples, y en todos los complejos, las acciones, espontáneas o reactivas, son causadas por órdenes que provienen de un cerebro. (Adviértase que ciertos organismos privados de cerebro, pero capaces de movimiento, precedieron a organismos que tienen cuerpo y cerebro, y después coexistieron con ellos).
No todas las acciones que ordena un cerebro son fruto de la deliberación. Por el contrario, se puede suponer que la mayoría de las acciones, teóricamente motivadas por el cerebro, que se desarrollan en este momento en el mundo, no son deliberadas en absoluto. Son respuestas simples, de las cuales un reflejo es buen ejemplo: un estímulo transportado por una neurona que incita a actuar a otra.
A medida que los organismos adquirieron mayor complejidad, las acciones «cerebro-motivadas» requirieron un proceso más mediatizado. Otras neuronas se interpolaron entre la neurona estimulante y la reactiva y de este modo se establecieron diversos circuitos paralelos, lo que no quiere decir que los organismos provistos de cerebros más complicados tuvieran por necesidad una mente. Los cerebros pueden tener muchos pasos intermedios en los circuitos interpolados entre el estímulo y la respuesta, y no por ello tienen mente; a menos que cumplan una condición esencial: la capacidad de desplegar imágenes internamente y de ordenarlas en un proceso llamado pensamiento. (Las imágenes no sólo son visuales; hay también «imágenes auditivas», «olfativas», y muchas otras). Ahora puedo completar mi afirmación acerca de los organismos conductuales diciendo que no todos tienen mente, esto es, no todos tienen fenómenos mentales (que es lo mismo que decir que carecen de cognición o de procesos cognitivos). Algunos organismos tienen cognición y comportamiento. Otros tienen acciones inteligentes, pero no mente. Ningún organismo parece tener mente pero no acción.
Mi posición, entonces, es que un organismo provisto de mente forma representaciones neurales que pueden transformarse en imágenes, manipularse en un proceso llamado pensamiento y finalmente influir en la conducta ayudando a predecir el futuro, a trazar planes conforme a esa previsión y a elegir la acción siguiente. Aquí está el quid de la neurobiología, tal como yo la imagino: el proceso por el cual representaciones neurales —consistentes en modificaciones biológicas derivadas del aprendizaje en un circuito neuronal— se transforman en imágenes en nuestra mente; el proceso que permite que cambios microestructurales invisibles en los circuitos neuronales (en los cuerpos celulares, dendritas y axones, y sinapsis), se conviertan en una representación neural que a su vez se transmuta en una imagen que cada uno experimenta como propia.
En una primera aproximación, la función global del cerebro es estar bien informado acerca de lo que sucede en el resto del cuerpo propiamente tal; acerca de lo que acaece en sí mismo; y acerca del entorno que rodea al organismo, para lograr así una acomodación adecuada y vivible entre organismo y medio ambiente. Desde una perspectiva evolutiva, no es al revés. Si no hubiera existido cuerpo, no habría habido cerebro. Por lo demás, los organismos simples, equipados con un cuerpo y comportamiento, pero sin cerebro ni mente, son, de hecho, mucho más numerosos que los humanos y en varios órdenes de magnitud. Basta pensar en las miríadas de felices bacterias —como las Escherichia coli— que viven en este instante dentro de cada uno de nosotros.
INTERACCIÓN DEL ORGANISMO Y EL MEDIO: ENFRENTAR EL MUNDO EXTERNO
Si cuerpo y cerebro interactúan intensamente, no menos lo hacen —por intermedio de su movimiento y dispositivos sensoriales— el organismo que conforman y su entorno.
El medio ambiente marca al organismo de diversas maneras. Una de ellas es la estimulación de la actividad neural en el ojo (dentro del cual está la retina), el oído (que contiene la cóclea, dispositivo detector de sonidos, y el vestíbulo, aparato que detecta el equilibrio), y las miríadas de terminales nerviosas en la piel, papilas gustativas y mucosas nasales. Las terminales nerviosas envían señales a determinados puntos de ingreso en el cerebro, conocidos como capas corticales sensoriales primarias de la visión, el oído, las sensaciones somáticas, el gusto y el olfato. Se las puede imaginar como especies de puertos a los cuales llegan las señales. Cada una de las regiones sensoriales primarias (capas corticales primarias visuales, auditivas, etc.) es una colección de varias áreas, y hay un denso entrecruzamiento de señales entre la agregación de áreas en cada colección sensorial primaria, como se puede apreciar en la figura 5-1. Más adelante, en este capítulo, propongo que estos sectores, íntimamente interconectados, son la base de representaciones topográficamente organizadas, la fuente de las imágenes mentales.

Figura 5-1. Diagrama simplificado de algunas interconexiones entre «capas corticales visuales primarias» (V1, V2, V3, V4, V5) y tres estructuras visuales subcorticales: núcleo geniculado lateral (LGN); el pulvinar (PUL) y el colliculus superior (coll). V1 también se conoce como corteza visual «primaria», y corresponde al área 17 de Brodmann. Nótese que la mayoría de los componentes de este sistema está interconectada por proyecciones neuronales aferentes y eferentes (flechas). La alimentación visual del sistema viene desde el ojo por intermedio de LGN y colliculus. Las entregas del sistema surgen en forma paralela de muchos de los componentes (i. e. de V4, V5 y así sucesivamente), hacia objetivos corticales y subcorticales.
A su vez, el organismo actúa sobre el entorno mediante movimientos de todo él cuerpo, miembros y aparato vocal, que están controlados por las capas corticales M 1, M 2 y M 3 (capas en las cuales también se generan los movimientos corporales internos) y la cooperación de varios núcleos motores subcorticales. Existen, por lo tanto, sectores del cerebro en que llegan continuamente señales del cuerpo propiamente tal o de los órganos sensoriales del cuerpo. Esos sectores de «entrada» (input) están anatómicamente separados y no se comunican de manera directa entre sí. También hay sectores cerebrales en los cuales surgen señales motoras y químicas; entre esos sectores de «salida» (output) están el tronco encefálico, los núcleos hipotalámicos y las capas corticales motoras.
UNA DIGRESIÓN SOBRE LA ARQUITECTURA DE LOS SISTEMAS NEURALES
Supongamos que tienes que inventar un cerebro humano empezando de la nada, y que has delineado los puertos a los cuales enviarás las múltiples señales sensoriales. ¿No sería deseable fusionar lo más rápidamente posible las señales que provienen de fuentes distintas —digamos, visión y audición— de manera que el cerebro generara «representaciones integradas» de las cosas que simultáneamente se ven y se oyen? ¿No querrías conectar dichas representaciones a controles motores para que el cerebro pudiera responder a ellas adecuadamente? Supongo que contestarás con un rotundo sí; pero esa no fue la respuesta de la naturaleza. Como muestra el estudio, decisivo, de las conexiones neuronales, que efectuaron hace un par de décadas E. G. Jones y T. P. S. Powell, la naturaleza no permite que los puertos sensoriales se comuniquen directamente, y tampoco deja que hablen directamente a los controles motores.[1] En el nivel de la corteza cerebral, por ejemplo, cada colección de áreas sensoriales primarias debe hablar primero a diversas regiones interpuestas, las que a su vez se comunican con zonas más distantes, y así sucesivamente. La conversación se realiza mediante axones proyectados hacia adelante, o proyecciones eferentes, que convergen en regiones río abajo que a su vez convergen en otras regiones.
Pareciera que esas múltiples corrientes paralelas y convergentes terminan en algunos puntos apicales, como la corteza cercana al hipocampo (corteza entorhinal) o algunos sectores de la corteza prefrontal (en las regiones dorsolateral o ventromedial). Sin embargo, esto no es totalmente exacto. En primer lugar, nunca «terminan» como tales, porque desde la vecindad de cada punto hacia el cual se proyectan adelante surge una proyección recíproca inversa hacia atrás. Es más apropiado decir que las señales se mueven corriente abajo y corriente arriba. En vez de una corriente hacia adelante, uno encuentra bucles de proyecciones hacia adelante y hacia atrás, que pueden crear una recurrencia perpetua.
Otra razón por la cual los flujos no «terminan» en sentido estricto es que, desde algunas de sus postas de relevo —especialmente las situadas más adelante— hay proyecciones directas hacia los controles motrices.
Así, la comunicación entre los sectores de entrada de información, y entre los sectores de entrada y salida, es indirecta, mediatizada, y utiliza una compleja arquitectura de agregados neuronales interconectados. A nivel de la corteza cerebral, esas agregaciones corresponden a regiones corticales situadas dentro de diversas cortezas asociadas. Pero la comunicación mediatizada también se produce por grandes núcleos subcorticales, como los que hay en el tálamo y en los ganglios basales, y por núcleos pequeños como los del tronco del cerebelo.
En fin, la cantidad de estructuras cerebrales situadas entre los sectores de entrada y salida es muy grande, e inmensa la complejidad de sus conexiones. La pregunta natural es: ¿Qué sucede en esas estructuras «interpuestas», de qué nos sirve toda esa complejidad? La respuesta es que esa actividad que hay allí, junto con la de las áreas de entrada y salida, construye puntualmente y manipula clandestinamente nuestras imágenes mentales. Sobre la base de esas imágenes (acerca de las cuales diré más en las páginas que siguen) interpretamos las señales que nos llegan a las capas corticales primarias, para organizarías en conceptos y categorizarlas. Podemos adquirir estrategias racionales y decisorias; y podemos seleccionar una respuesta motora en el menú disponible en el cerebro, o formular una nueva reacción motora, un conjunto de acciones deliberadas y voluntarias que pueden ir desde golpear una mesa hasta acariciar un niño, escribir una carta al editor o tocar a Mozart en el piano.
Entre los cinco grandes sectores sensoriales de entrada y los tres principales de salida, están las capas corticales asociativas, los ganglios basales, el tálamo, las capas corticales y los núcleos del sistema límbico, el tronco del encéfalo y el cerebelo. En conjunto, este «órgano» de información y de gobierno, esta gran colección de sistemas, guarda conocimientos adquiridos e innatos sobre el cuerpo propiamente tal, el mundo externo y el cerebro mismo en cuanto interactúa con el cuerpo propiamente tal y con el mundo externo. Este conocimiento sirve para desplegar y manipular producciones motoras y mentales, las imágenes que constituyen nuestro pensamiento. Creo que este depósito de hechos y de estrategias para su manipulación está almacenado, latente y en suspenso, como «representaciones disposicionales» (para abreviar: «disposiciones»), en los sectores intersticiales del cerebro. Así, la regulación biológica, la memoria de estados previos y la planificación de acciones futuras no sólo resultan de la actividad cooperativa de las capas corticales primarias motoras y sensoriales, sino también de los sectores intersticiales.
UNA MENTE INTEGRADA A PARTIR DE ACTIVIDADES DISPERSAS
Muchos de quienes gozan pensando cómo trabaja el cerebro comparten la intuición —falsa— de que las distintas vetas de procesamiento sensorial que se experimentan en la mente —vistas y sonidos, sabor y aroma, textura y forma— «suceden» en una estructura cerebral única. Parecería razonable pensar que lo que está junto en la mente también lo está en un solo lugar del cerebro donde se mezclarían diferentes aspectos sensoriales. La metáfora habitual semeja una gran pantalla de cinemascope, equipada para proyectar en glorioso tecnicolor, sonido estereofónico, y que quizás también posee una banda de aromas. Sobre esta concepción, a la que llama «teatro cartesiano», Daniel Dennett ha escrito en detalle y argumentado persuasivamente —sobre bases cognitivas— que ese escenario cartesiano no puede existir.[2] Apoyándome en fundamentos neurocientíficos, yo también sostengo que la intuición es falsa.
Resumo ahora mis razones, que ya he expuesto detenidamente en otro trabajo.[3] Contra la idea de un sector cerebral integrado, argumento que ninguna región del cerebro humano está equipada para procesar simultáneamente representaciones sensoriales de todas las modalidades activas cuando experimentamos sincrónicamente —sonido, movimiento, forma y color, por ejemplo— en un registro temporal y espacial perfecto.
Estamos empezando a indagar dónde es probable que se construyan las imágenes correspondientes a cada diferente modalidad, pero en ninguna parte hemos encontrado una zona única hacia la cual todos esos distintos productos se proyecten en un registro exacto.
Es verdad que hay pocas regiones cerebrales donde puedan converger señales provenientes de muchas áreas sensoriales primarias diferentes. Algunas de esas zonas de convergencia, como las capas corticales entorhinales y perihinales, reciben una amplia variedad de señales polimodales. Pero no es probable que el tipo de integración que pueden producir esas regiones utilizando esas señales sea el que forme la base de una mente integrada. Desde luego, una lesión en esas zonas superiores de convergencia no excluye en absoluto —aun cuando ocurra en ambos hemisferios— la integración de la «mente», aunque cause otras consecuencias neurosicológicas (como dificultades de aprendizaje, por ejemplo).
Quizás sea más fructífero pensar que nuestra sensación de integración mental deriva de la acción concertada de sistemas de gran escala, que sincronizan conjuntos de actividad neural en distintas regiones cerebrales. Si la actividad ocurre casi al mismo tiempo en zonas cerebrales anatómicamente separadas, es posible unir los cuadros detrás de la escena y crear la impresión de que todo sucede en el mismo lugar. Nótese que de ninguna manera es ésta una explicación de cómo el tiempo establece la vinculación, sino más bien una sugerencia de que la cadencia secuencial es parte importante del mecanismo. La idea de la integración tiempo mediante se ha presentado en la década pasada, y varios teóricos la usan de manera muy destacada en sus trabajos.[4]
Que el cerebro integre procesos separados en combinaciones significativas mediante el tiempo es una solución económica y sensata, pero presenta riesgos y problemas. El mayor riesgo es la asincronía. Cualquier dificultad en el mecanismo rítmico secuencial podría crear una integración espuria, o una desintegración. Es lo que muy probablemente acontece en los estados de confusión que suceden a lesiones craneanas, o en algunos síntomas esquizofrénicos y otros males. El problema fundamental que crea la vinculación temporal se relaciona con la necesidad de mantener una actividad focalizada en diferentes sitios durante el tiempo necesario para construir combinaciones significativas y para efectuar razonamientos y adoptar decisiones. En otras palabras, la vinculación temporal requiere de mecanismos poderosos y efectivos de atención y memoria operativa, y parece que la naturaleza está dispuesta a proporcionarlos.
Cada sistema sensorial parece estar equipado para proveer sus propios instrumentos de atención local y memoria operativa. Pero cuando se trata de los procesos de atención global y memoria operativa, los estudios experimentales en animales y en humanos sugieren que son esenciales las capas corticales prefrontales y algunas estructuras del sistema límbico (la corteza cingular anterior).[5] La misteriosa conexión entre los procesos y sistemas cerebrales, expuesta a comienzos de este capítulo, puede estar más clara ahora.
IMÁGENES ACTUALES, IMÁGENES DEL PASADO, IMÁGENES DEL FUTURO
El conocimiento fáctico que se requiere para razonar y tomar decisiones llega a la mente con la forma de imágenes. Examinemos brevemente los probables sustratos neurales de esas imágenes.
Si contemplamos el paisaje otoñal, o escuchamos la música que resuena a lo lejos, o rozamos con los dedos una lisa superficie de metal, o leemos estas palabras línea por línea recorriendo esta página, estamos percibiendo y por ende formando imágenes de diversas modalidades sensoriales. Las imágenes así formadas se llaman imágenes perceptuales.
Pero podemos dejar de prestar atención al paisaje, música, superficie o texto, distraernos y dirigir los pensamientos a otra parte. Quizás pensemos en la tía Julia, en la Torre Eiffel, en la voz de Plácido Domingo o en lo que acabo de decir de las imágenes. Independientemente de que estén hechos de formas, colores, movimientos, tonalidades, palabras verbalizadas o no, esos pensamientos también están constituidos por imágenes. Estas, que ocurren cuando recuperamos un recuerdo de cosas pasadas, son llamadas imágenes evocadas, para distinguirlas de la variedad perceptual.
Por intermedio de las imágenes evocadas, podemos recuperar un tipo particular de imagen pasada, una que formamos al planear algo que aún no ha sucedido pero que queremos que suceda, como, por ejemplo, reorganizar la biblioteca este fin de semana. A medida que se despliega el proceso de planeamiento, vamos formando imágenes de objetos y movimientos y consolidando una memoria de esa ficción en la mente. Las imágenes de algo que aún no sucede, y que de hecho puede no ocurrir nunca, no poseen una naturaleza diferente de las imágenes que tenemos de algo que sí ha sucedido. Antes que recuerdo del pasado que fue, son la memoria de un futuro posible.
Estas diversas imágenes —perceptuales, evocadas del pasado efectivo y evocadas de planes del futuro— son construcciones del cerebro de nuestro organismo. Todo lo que podemos saber con certeza es que son reales para nosotros mismos, y que otros seres fabrican imágenes comparables. Compartimos con los otros humanos —e incluso con algunos animales— ese concepto basado en imágenes del mundo; hay una notoria coherencia en las construcciones que hacen distintos individuos de los aspectos esenciales del entorno (texturas, sonidos, formas, colores, espacio). Si nuestros organismos estuvieran diseñados de otra manera, las construcciones que hacemos del mundo que nos rodea serían diferentes. No sabemos, y no es probable que algún día lleguemos a saber, cómo es la realidad «absoluta».
¿Cómo llegamos a crear estas maravillosas construcciones? Pareciera que son confeccionadas por una compleja maquinaria neural de percepción, memoria y razonamiento. A veces la construcción se extrae del mundo exterior al cerebro —esto es, del mundo que está adentro y alrededor de nuestro cuerpo— con alguna ayuda de la memoria pasada. Así sucede cuando generamos imágenes perceptuales. Otras, la construcción es dirigida desde el interior del cerebro, mediante nuestro suave y silencioso proceso pensante, de arriba hacia abajo, por decirlo así. Ocurre, por ejemplo, cuando recordamos una melodía o visualizamos una escena con los ojos cerrados sin que importe si repite un acontecimiento real o uno imaginario.
Pero la actividad neural que se relaciona de modo más íntimo con las imágenes que experimentamos, ocurre en la corteza sensorial primaria y no en otras regiones. Mediante la percepción o por intermedio del recuerdo, la actividad de esa corteza resulta, por decirlo así, de procesos complejos que operan tras bambalinas en numerosas regiones de la corteza cerebral y de los núcleos neuronales bajo ésta, en el tallo cerebral, ganglios basales y otras zonas. En breve: las imágenes se basan directamente en aquellas representaciones neurales —y sólo en aquellas— que están topográficamente organizadas y que ocurren en las capas corticales sensoriales primarias. Pero se forman o bien bajo el control de receptores sensoriales orientados hacia el exterior del cerebro (la retina, por ejemplo), o bien bajo el control de representaciones disposicionales (disposiciones) contenidas en el cerebro, en regiones corticales y en núcleos subcorticales.
FORMACIÓN DE IMÁGENES PERCEPTUALES
¿Cómo se forman las imágenes cuando estamos percibiendo algo en el mundo (un paisaje, por ejemplo) o en el cuerpo (un dolor en el codo derecho)? En ambos casos, un primer paso es necesario, pero no suficiente: señales del sector corporal correspondiente (ojo y retina, o bien terminales nerviosas en la articulación del codo) son transportadas hasta el cerebro por las neuronas, a lo largo de sus axones y a través de varias sinapsis electroquímicas. Las señales son entregadas a las capas corticales primarias[1*]. En el caso de las señales desde la retina, la entrega se verifica en las capas corticales primarias visuales, situadas atrás en el cerebro, en el lóbulo occipital. En el caso de las provenientes de la articulación del codo, la entrega se produce en las capas corticales primarias somatosensoriales de las regiones parietal e insular, parte del sector cerebral que se daña en la anosognosia. Nótese, otra vez, que ésta es una colección de áreas y no un solo centro. Las áreas que forman la colección son individualmente complejas, y la malla de interconexiones que integran lo es aún más. Las representaciones topográficamente organizadas resultan de la interacción concertada de esas áreas y no de la operación aislada de una de ellas. Esta idea no tiene nada de frenológico.
Cuando son destruidas todas o la mayoría de las capas corticales sensoriales primarias de una modalidad determinada, desaparece la capacidad para formar imágenes en ese modo específico. Poco pueden ver los pacientes privados de capas primarias visuales. (Un quantum de capacidad sensorial residual permanece, probablemente, porque las estructuras corticales y subcorticales relacionadas con esa modalidad están intactas. Después de una extensa destrucción en las capas corticales primarias visuales, algunos pacientes pueden señalar puntos luminosos que dicen no ver; tienen lo que se conoce como «visión ciega». Las capas corticales parietales, el tubérculo cuadrigémino superior y el tálamo son algunas de las estructuras posiblemente implicadas en esos procesos). El defecto perceptual puede ser muy específico. Después del daño de uno de los subsistemas en las capas corticales visuales primarias, por ejemplo, se puede perder la capacidad de percibir colores: la pérdida puede ser total, o una atenuación (caso en el cual los pacientes ven colores desvaídos). Los afectados ven formas, movimientos y profundidad, pero no color. En esta situación, conocida como acromatopsia, construyen el universo en diversas tonalidades de gris.
Aunque las capas sensoriales primarias y las representaciones topográficamente organizadas que construyen sean necesarias para que las imágenes acontezcan en la conciencia, parecen, no obstante, ser insuficientes. Dicho de otra manera: dudo mucho que fuéramos conscientes de imagen alguna si nuestro cerebro sólo generara finas representaciones topográficamente organizadas y no hiciera nada más con ellas. ¿Cómo sabríamos que son nuestras imágenes? La subjetividad, clave de la conciencia, faltaría en el diseño. Deben cumplirse otras condiciones.
En esencia, esas representaciones neurales deben ser correlativas con aquellas que, momento a momento, constituyen la base neural del self (sí mismo). Esta cuestión surgirá nuevamente en los capítulos 7 y 10, pero digamos desde ya que el self no es el homúnculo infame, un enano dentro de nuestro cerebro, que percibe y piensa con las imágenes que el cerebro construye. Es, más bien, un estado neurobiológico perpetuamente recreado. Años de justificados ataques al concepto del homúnculo hacen que muchos teóricos se muestren igualmente temerosos del self. Pero el self neural no necesita en absoluto ser homuncular. Lo que debería causar miedo, en realidad, es la idea de una cognición privada de self.
ALMACENAMIENTO DE IMÁGENES Y FORMACIÓN DE IMÁGENES EVOCABLES
Las imágenes no se almacenan como facsímiles de cosas, acontecimientos, palabras o frases. El cerebro no archiva fotografías Polaroid de gentes, objetos, paisajes; tampoco acopia cintas de audio, musicales o discursivas; no acumula películas de escenas de nuestra vida; ni guarda tarjetas con apuntes o transparencias Teleprompters, como las que ayudan a los políticos a ganarse el pan de cada día. En suma, parece que de nada existen estampas, aun miniaturizadas; no hay microfilmes ni microfichas; no hay copias en disco duro. En vista del enorme bagaje de conocimientos que adquirimos a lo largo de la vida, cualquier tipo de almacenamiento facsimilar plantearía problemas insuperables de capacidad. Si el cerebro fuera como una biblioteca, nos quedaríamos sin estantes, igual que sucede en las bibliotecas. Además, con el acopio de facsímiles, la recuperación de información se tornaría problemática. Tenemos evidencia directa de que cuando recordamos un objeto determinado, una cara, una escena, no logramos una reproducción exacta del original, sino más bien una interpretación, una versión nueva, reconstruida. También, a medida que avanzamos en años y cambia nuestra experiencia, las versiones evolucionan. Nada de ello es compatible con una representación facsimilar rígida, como indicó el psicólogo inglés Frederic Bartlett hace algunas décadas, cuando postuló que la memoria es esencialmente reconstructiva.[7]
Sin embargo, negar que puedan existir estampas en el cerebro debe reconciliarse con la sensación —que todos compartimos— de que podemos evocar en la mente, ojos y oídos, aproximaciones de imágenes pasadas. El que dichas aproximaciones no sean precisas, o que sean menos vividas que los originales que reproducen, no contradice este hecho.
Una respuesta provisional sugiere que esas imágenes mentales son construcciones puntuales, intentos de replicar patrones alguna vez experimentados; si bien la probabilidad de una reproducción exacta es baja, la eventualidad de una réplica sustancial es mayor o menor según las circunstancias en que las imágenes fueron aprendidas o estén siendo evocadas. Estas imágenes evocadas, que tienden a permanecer fugazmente en la conciencia, aunque parecen buenas réplicas, suelen ser imprecisas o incompletas. Sospecho que las imágenes mentales evocadas explícitas surgen de la activación transitoria y sincrónica de patrones neurales de descarga, principalmente en las mismas capas sensoriales primarias donde ocurrieron en su momento los patrones de activación correspondientes a representaciones perceptuales. De la activación resulta una representación topográficamente organizada.
Varios argumentos y alguna evidencia favorecen esta noción. En la condición conocida como acromatopsia, ya descrita, el daño local en las capas corticales visuales primarias no sólo deteriora la percepción sino la imaginería cromática. Si eres acromatópsico no puedes imaginar el color. Si te pido que imagines una banana, podrás figurarte su forma pero no su colorido; lo verás en tonos de gris. Si el «conocimiento cromático» estuviera almacenado en otra parte, en un sistema separado del que mantiene la «percepción cromática», los pacientes acromatópsicos podrían imaginar los colores, aunque no fueran capaces de percibirlos en los objetos externos. Pero no pueden.
Los pacientes con daño extenso en las capas corticales visuales primarias pierden la capacidad de generar imaginería visual. No obstante, pueden recordar las propiedades táctiles y espaciales de los objetos, así como las imágenes sonoras.
Los estudios preliminares de la evocación visual, efectuados con la Tomografía por Emisión de Positrones (PET) —una técnica de neuroimaginería— y con la Resonancia Magnética Funcional (FMR), respaldan esta idea. Steven Kosslyn y su grupo, y Hanna Damasio, Thomas Grabowski y sus colegas, han descubierto que la evocación de imágenes visuales activa, entre otras áreas, las capas corticales visuales primarias.[8]
¿Cómo damos forma a las representaciones-topográficamente organizadas que son necesarias para experimentar imágenes evocadas? Pienso que esas representaciones se construyen momento a momento por orden de patrones neurales disposicionales adquiridos, y localizados en otras partes del cerebro. Uso ese término porque lo que hacen, literalmente, es accionar otros patrones neurales, generar actividad neuronal en otros lugares del cerebro, en circuitos que forman parte del mismo sistema y con los que hay abundante interconexión neuronal. Los moldes disposicionales existen como patrones potenciales de actividad neuronal en pequeños conjuntos de neuronas que llamo «zonas de convergencia»; esto es, en una serie de neuronas que activan disposiciones en el conjunto. Las disposiciones relativas a imágenes evocables se adquirieron mediante aprendizaje, y así podemos decir que constituyen una memoria. Las zonas de convergencia cuyas representaciones disposicionales pueden traducirse en imágenes cuando retroestimulan las capas corticales sensoriales primarias, están distribuidas en las capas corticales superiores de asociación (en las regiones occipital, temporal, parietal y frontal), así como en los ganglios basales y en las estructuras límbicas.
Las representaciones disposicionales no almacenan un cuadro propiamente tal en su pequeña comunidad de sinapsis, sino los medios para reconstituir «un cuadro». Si tienes una representación disposicional del rostro de tía Margarita, esa representación no contiene su cara como tal, sino los patrones de descarga que gatillan la reconstrucción momentánea de una representación aproximada del rostro de tía Margarita en las capas corticales visuales primarias.
Para que el semblante de tía Margarita aparezca en las pantallas de tu mente, deben retroencenderse —de manera más o menos sincrónica— diferentes representaciones disposicionales situadas en varias capas corticales visuales y de asociación superior (sospecho que principalmente en las regiones temporal y occipital).[9] El mismo orden debería ocurrir en el terreno auditivo. En las capas corticales de asociación auditiva hay representaciones disposicionales de la voz de tía Margarita, que pueden retroactivar las capas corticales auditivas primarias y generar momentáneamente una representación aproximada de la voz de tía Margarita.
No hay una sola fórmula oculta para esta reconstrucción. Tía Margarita, como persona completa, no existe en un lugar único de tu cerebro. Está diseminada por todo él, en multitud de representaciones disposicionales de esto y aquello. Y cuando evocas remembranzas de aspectos Margarita, y ella reaparece como representaciones topográficas en varias capas corticales primarias (visuales, auditivas, y otras), sólo se hace presente en forma de vistas separadas durante el lapso en que construyes algún significado de su persona.
Si dentro de cincuenta años, en un experimento imaginario, te sumergieras en las representaciones disposicionales visuales que un tercero tuviera de tía Margarita, puedo predecir desde ya que no verías nada que se pareciera efectivamente al rostro de tía Margarita, porque las representaciones disposicionales no están topográficamente organizadas. Pero, si inspeccionaras los patrones de actividad en las capas corticales visuales primarias de ese tercero, dentro de los cien primeros milisegundos posteriores al retrogatillaje del rostro de tía Margarita en la zona de convergencia, probablemente detectarías algunos patrones de actividad que tuvieran cierta relación con la geografía del rostro de tía Margarita. Habría alguna correspondencia entre lo que sabías de su cara y el patrón de actividad en los circuitos de las capas corticales primarias visuales de ese tercero que también la conoció y que en ese momento piensa en ella.
Ya hay indicios probatorios en este sentido. R. B. H. Tootell, mediante un método de imágenes neuroanatómicas, ha demostrado que cuando un mono percibe ciertas formas —como una cruz o un cuadrado—, la actividad neuronal en las capas corticales visuales primarias se organiza topográficamente en una pauta que replica las formas que el mono está viendo.[10] Dicho en otras palabras: un observador independiente, al mirar el estímulo externo y los patrones de actividad neuronal, reconocerá una semejanza estructural. (Ver Fig. 5-2). El mismo razonamiento es válido para el hallazgo de Michael Merzenich acerca de patrones dinámicos de representación corporal en las capas corticales somatosensoriales.[11] Conviene advertir, sin embargo, como ya señalé anteriormente, que tener dicha representación en la corteza cerebral no es equivalente a tener conciencia de ella. Es necesario, pero no suficiente.
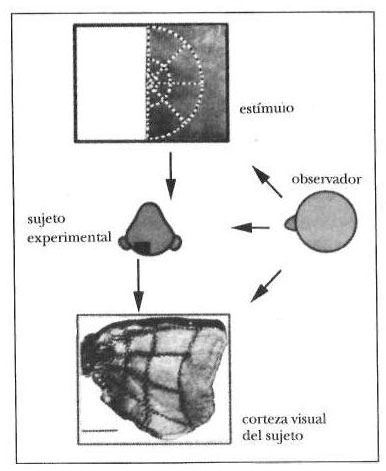
Figura 5-2. Un observador, que mirara el estímulo presentado a un animal sujeto de un experimento, y que después examinara la activación motivada por ese estímulo en el córtex visual del animal, descubriría una coherencia notable entre la forma del estímulo y la del patrón de actividad neural en una de las capas de la corteza visual primaria (Capa 4C). El estímulo y la imagen cerebral provienen de los trabajos de Roger Tootell, que realizó este experimento.
Estoy llamando representación disposicional a una potencialidad latente de encendido que se activa cuando las neuronas descargan, conforme a un patrón determinado, con un ritmo, duración y secuencia precisos y un objetivo específico que resulta ser otro conjunto de neuronas. Nadie sabe cómo son los «códigos» comprendidos en el conjunto, a pesar de la multitud de descubrimientos recientes que se han acumulado en el estudio de la modificación sináptica. Sin embargo, esto parece probable: los patrones de descarga resultan del fortalecimiento o debilitamiento de las sinapsis, lo que a su vez es consecuencia de los cambios funcionales que ocurren a nivel microscópico en las ramas fibrosas (axones y dendritas) de las neuronas.[12]
Las representaciones disposicionales existen en estado potencial, susceptibles de activación, como el poblado de Brigadoon[2*].
EL CONOCIMIENTO ESTA INCORPORADO EN LAS REPRESENTACIONES DISPOSICIONALES
Las representaciones disposicionales son el depósito total de nuestros conocimientos; abarcan tanto el conocimiento innato como el adquirido mediante la experiencia. El conocimiento innato se basa en representaciones disposicionales situadas en el hipotálamo, en el tallo cerebral y en el sistema límbico. Puedes conceptualizarlo como mandatos relativos a la regulación biológica, necesarios para la supervivencia (por ejemplo, los mandatos que controlan el metabolismo, las pulsiones y los instintos). Controlan diversos procesos, pero por lo general no se transmutan en imágenes mentales. Hablaré de ellos en el próximo capítulo.
Los conocimientos adquiridos se basan en representaciones disposicionales situadas en capas corticales superiores y en numerosos núcleos de materia gris situados debajo del nivel cortical. Algunas de estas representaciones disposicionales contienen registros del conocimiento convertible en imágenes que podemos evocar y que utilizamos para movernos, razonar, la creatividad y la planificación; y algunas contienen registros de normas y estrategias con las cuales operamos dichas imágenes. La adquisición de conocimiento nuevo se logra mediante la continua modificación de esas representaciones disposicionales.
Cuando se activan las representaciones disposicionales puede haber distintos resultados. Pueden gatillar otras representaciones disposicionales con las cuales estén íntimamente relacionadas por el diseño del circuito (por ejemplo, las representaciones disposicionales de la corteza temporal, podrían encender a sus homologas en la corteza occipital, que forman parte de los mismos sistemas reforzados). O pueden generar una representación topográficamente organizada gatillando directamente hacia capas corticales sensoriales primarias o activando otras representaciones disposicionales en el mismo sistema reforzado. O pueden generar un movimiento al activar una corteza motriz o núcleo como los ganglios basales.
La aparición de una imagen evocada resulta de la reconstrucción de un patrón transitorio (metafóricamente, un mapa) en las capas corticales sensoriales primarias, reconstitución gatillada por la activación de representaciones disposicionales en otras zonas del cerebro (como en las cortezas asociativas). El mismo tipo de activación «cartográfica» ocurre en las capas corticales motoras, y es la base del movimiento. Las representaciones disposicionales en que se apoya el movimiento están situadas en las capas corticales premotoras, los ganglios basales y las capas corticales límbicas. Hay indicios de que activan tanto los movimientos como las imágenes internas de los movimientos corporales; debido a la naturaleza veloz de los movimientos, estas últimas suelen quedar encubiertas, en la conciencia, por nuestra percepción del movimiento mismo.
EL PENSAMIENTO ESTÁ HECHO PRINCIPALMENTE DE IMÁGENES
Frecuentemente se afirma que el pensamiento no sólo está hecho de imágenes, sino también de palabras y de símbolos abstractos de índole no imaginaria. Nadie negará, por cierto, que el pensamiento incluye palabras y símbolos arbitrarios. Sin embargo, lo que falta en ese aserto, es que tanto las palabras como los símbolos arbitrarios se basan en representaciones topográficamente organizadas, pasibles de convertirse en imágenes. La mayoría de las palabras que usamos en nuestro discurso interno, antes de hablar o escribir, existe en nuestra consciencia en forma de imágenes auditivas o visuales. Si no se transformaran en imágenes —aún fugaces— no podrían convertirse en algo conocible.[13] Esto es válido incluso para aquellas representaciones topográficamente organizadas que no se muestran en la luz de nuestra consciencia, sino que se activan encubiertamente. Gracias a experimentos de incitación preparatoria sabemos que, si bien esas representaciones son elaboradas de manera subrepticia, pueden influir el curso del proceso de pensamiento, e incluso aparecer inesperadamente en la consciencia un poco después. (La incitación preparatoria consiste en activar una representación en forma incompleta, o activarla sin prestarle atención).
Experimentamos habitualmente este fenómeno. Después de una animada conversación entre varias personas, una palabra o un aserto que no escuchamos durante la charla nos aparece de súbito en la mente. Quizá nos sorprenda el no haberlo escuchado e incluso dudemos de su realidad, preguntando por ejemplo, «¿dijiste tal y tal cosa?». Por cierto X dijo tal y tal cosa, pero, como estabas concentrado en lo que decía Y, no prestaste atención a las representaciones cartográficas que se formaron en relación a lo que decía X, y de ellas sólo quedó una memoria disposicional. Cuando se relajó tu concentración en Y —y si la palabra o frase elidida era importante para ti— las representaciones disposicionales reconstruyeron una representación topográficamente organizada en alguna capa cortical sensorial primaria que se convirtió en una imagen, pues eras consciente de ella. Por lo demás, no habrías formado jamás una representación disposicional sin fabricar primero una representación perceptual topográficamente cartografiada: parece no existir una vía anatómica para liberar una información sensorial compleja en las capas corticales asociativas que sostienen las representaciones disposicionales sin antes detenerse en las capas corticales primarias. (Es posible que esto no sea válido en el caso de informaciones sensoriales no complejas).
Estos comentarios también son aplicables a los símbolos que podemos usar en la solución mental de un problema matemático (si bien quizá no valgan para todas las formas de pensamiento matemático). Si esos símbolos no pudieran transformarse en imágenes no los conoceríamos, y seríamos incapaces de manipularlos conscientemente. En este sentido, es interesante observar que algunos matemáticos y físicos perspicaces dicen que su pensamiento está dominado por imágenes. Frecuentemente se trata de imágenes visuales, e incluso pueden ser somatosensoriales. No es sorprendente que Benoit Mandelbrot, cuyo trabajo de toda la vida es la geometría fractal, diga que siempre piensa en imágenes.[14] Cuenta que al físico Richard Feynman no le gustaba mirar una ecuación si antes no examinaba la ilustración adjunta (y nótese que tanto la ecuación como la ilustración son, de hecho, imágenes). En cuanto a Albert Einstein, no tenía duda alguna acerca de este proceso:
Las palabras o el lenguaje, tal como son escritos o hablados, no parecen jugar papel alguno en mi mecanismo de pensamiento. Las entidades psíquicas que parecen servir de elementos en el pensamiento son ciertos signos e imágenes más o menos claras que se pueden combinar y reproducir «voluntariamente». Por supuesto, hay alguna conexión entre esos elementos y conceptos lógicos relevantes. También es claro que el deseo de llegar al fin a conceptos lógicamente interconectados es la base emocional de este juego más bien vago con los elementos arriba mencionados.
Más adelante, en el mismo texto, aclara aún más su idea:
Los elementos arriba citados son, en mi caso, de tipo visual y muscular. Hay que buscar laboriosamente palabras convencionales y otros signos sólo en una etapa secundaria, cuando el juego asociativo descrito está suficientemente consolidado y se lo puede reproducir a voluntad.[15]
El punto es, entonces, que las imágenes son probablemente el contenido principal de nuestros pensamientos, con independencia de la modalidad sensorial en que se generan y sin que importe si se refieren a una cosa o a un proceso que involucra cosas; ni si están en un idioma determinado; ni si se refieren a palabras u otros símbolos que correspondan a una cosa o a un proceso. Ocultos detrás de esas imágenes, pocas veces o jamás conocidos por nosotros, hay numerosos procesos que guían la generación y despliegue de esas imágenes en el espacio y en el tiempo. Esos procesos utilizan normas y estrategias almacenadas en representaciones disposicionales. Son esenciales para nuestro pensar pero no son un contenido del pensamiento.
Las imágenes que reconstituimos mediante la evocación se presentan conjuntamente con las que se forman por intermedio de estímulos externos. Las imágenes recompuestas desde el interior del cerebro son menos nítidas que las incitadas desde el exterior. Son «débiles», como dice David Hume, en comparación con las «vividas» imágenes que generan los estímulos externos al cerebro. Pero, de todos modos, son imágenes.
ALGUNAS PALABRAS SOBRE DESARROLLO NEURAL
Tal como expuse antes, tanto los sistemas como los circuitos cerebrales, así como las operaciones que realizan, dependen del patrón de conexiones entre neuronas y de la fuerza de las sinapsis que constituyen esas conexiones. ¿Pero cómo se nos establecen esos patrones de conexión y las intensidades sinápticas en el cerebro? ¿Y cuándo? ¿Se apostan simultáneamente en todos los sistemas en todo el cerebro? Una vez establecidos, ¿quedan emplazados para siempre? Aún no hay respuestas definitivas para estas preguntas. A pesar de que los conocimientos sobre el tema fluyen constantemente, y nada puede darse por sentado, es probable que las cosas ocurran de este modo:
- El genoma humano (la suma total de los genes en nuestros cromosomas) no especifica la estructura completa del cerebro. No hay suficientes genes disponibles para determinar la exacta estructura y lugar de cuanto hay en nuestro organismo, especialmente en el cerebro, donde miles de millones de neuronas forman sus cornados sinápticos. La desproporción es enorme: contamos con aproximadamente 100.000 genes (105), pero más de 1.000.000.000.000.000 (mil billones, o 1015) de sinapsis en el cerebro. Además, la formación de tejidos genéticamente inducida es asistida por la interacción celular, en la cual moléculas celulares y substraíales de adhesión juegan un papel importante. Las interacciones de este tipo, que suceden en las células mientras se verifica el desarrollo celular temprano, controlan de hecho, en parte, la expresión de los genes que regulan dicho desarrollo. Hasta donde sabemos, entonces, numerosas características estructurales específicas están determinadas por los genes, en tanto que otro número importante puede estar determinado sólo por la actividad del organismo mismo mientras crece y continuamente se modifica a lo largo de su vida.[16]
- El genoma ayuda a establecer la estructura —precisa o casi precisa— de una multitud de circuitos y sistemas importantes en los sectores evolutivos arcaicos del cerebro humano. Aunque necesitamos con urgencia estudios modernos relativos al desarrollo de esos sectores cerebrales, y aunque muchas cosas podrían cambiar cuando dichos estudios se materialicen, la afirmación anterior parece razonablemente válida para el tallo cerebral, el hipotálamo, prosencéfalo basal, y especialmente para la amígdala y la región cingular. (Hablaré más detenidamente sobre esas estructuras y sus funciones en los siguientes capítulos). Compartimos lo esencial de esas regiones cerebrales con individuos de muchas otras especies. El rol principal de esas estructuras zonales es la regulación de los procesos vitales básicos, sin recurso a la mente y la razón. En esos circuitos, los patrones innatos[3*] de actividad neuronal no generan imágenes (aunque las consecuencias de su actividad pueden ser traducidas en imágenes); regulan mecanismos homeostáticos sin los cuales la supervivencia es imposible. Sin los circuitos innatamente instalados en esas áreas cerebrales, seríamos incapaces de respirar, regular los latidos cardíacos, equilibrar nuestro metabolismo, buscar alimento y refugio, evitar a los depredadores, y reproducirnos. Si careciéramos de esta regulación biológica prefabricada, cesaría la supervivencia individual y evolutiva. En fin, esos circuitos innatos tienen un rol que debo destacar porque habitualmente es ignorado cuando se conceptualizan las estructuras neurales que sostienen mente y conducta: Los circuitos innatos no sólo intervienen en la regulación biológica del cuerpo, sino también en el desarrollo y actividad adulta de las estructuras evolutivamente modernas del cerebro.
- Al resto del cerebro, ciertas estructuras —cuyas especificaciones son equivalentes a las que los genes instalan en los circuitos del tallo cerebral o del hipotálamo— llegan mucho después del nacimiento, a medida que el individuo se desarrolla a través de infancia, niñez y adolescencia y mientras interactúa con el entorno y con otros individuos. Se puede conjeturar con mucha certidumbre —por lo menos en lo que concierne a las áreas evolutivamente modernas del cerebro— que el genoma ayuda a establecer un ordenamiento más bien global, no detallado, de los sistemas y circuitos. ¿Pero cómo se produce el ordenamiento exacto? Bajo la influencia de circunstancias externas, complementadas y delimitadas por el influjo de los circuitos relativos a la regulación biológica, innatos y precisamente establecidos.
En pocas palabras, la actividad de los circuitos en los sectores del cerebro modernos y movidos por la experiencia (la neocorteza, por ejemplo) es indispensable para producir una clase particular de representaciones neurales en que se fundan la mente (imágenes) y las acciones conscientes. Pero la neocorteza no puede generar imágenes si el anticuado subterráneo del cerebro (hipotálamo, tallo cerebral) no está intacto y no coopera.
Este ordenamiento nos puede proporcionar una pausa. Hete aquí que tenemos circuitos innatos cuya misión es regular las funciones corporales y asegurar la supervivencia del organismo, lo que se logra mediante el control de las operaciones bioquímicas internas del sistema endocrino, del sistema inmune, de las vísceras, de las pulsiones y de los instintos. ¿Por qué deberían estos circuitos interferir con la formación de otros, más modernos y plásticos, dedicados a la representación de nuestras experiencias adquiridas? La respuesta a esta importante pregunta es que tanto los registros de experiencias como sus respuestas —si deben ser adaptativas— deben ser evaluadas y configuradas por un conjunto fundamental de preferencias del organismo que considera que la supervivencia es prioritaria. Como esta evaluación y configuración son vitales para la continuación del organismo, parece que los genes especificaran, además, que los circuitos innatos deben ejercer una influencia profunda en casi todo el conjunto de circuitos que puede ser modificado por la experiencia. Ese influjo lo realizan en gran medida neuronas «moduladoras», que actúan sobre el resto de los circuitos. Estas neuronas moduladoras se encuentran en el tallo cerebral y en el prosencéfalo basal, y son influidas en todo momento por las interacciones del organismo. Las neuronas moduladoras distribuyen neurotransmisores (como la dopamina, la norepinefrina, la serotonina y la acetilcolina) a amplias regiones de la corteza cerebral y de los núcleos subcorticales. Este hábil arreglo puede ser descrito como sigue: 1) los circuitos innatos regulatorios están involucrados en la tarea de asegurar la supervivencia del organismo y, por ello, están secretamente informados de lo que sucede en regiones más modernas del cerebro; 2) lo bueno y lo malo de las situaciones que el organismo enfrenta les son señalados continuamente, y 3) expresan su intrínseca reacción a esas características influyendo en la configuración del resto del cerebro, de manera que este contribuya del modo más eficaz a la supervivencia.
Así, a medida que nos desarrollamos desde la infancia hacia la edad adulta, el diseño de los circuitos cerebrales que representa nuestro cuerpo en evolución y su interacción con el mundo parece depender de las actividades en las que el organismo se involucra, y de la acción de la circuitería biorreguladora innata según como esta última reaccione frente a esas actividades. Esta descripción pone de manifiesto lo inadecuado que es concebir cerebro, conducta y mente en términos de «naturaleza versus crianza» o de genes vs. experiencia. Ni el cerebro ni la mente son tabulae rasae cuando nacemos. Sin embargo, tampoco están genéticamente determinados en su totalidad. La sombra genética es grande, pero incompleta. Los genes proveen una estructura precisa a uno de los componentes cerebrales; en el otro, la estructura precisa está por ser determinada. La estructura por determinar sólo puede lograrse bajo el influjo de tres elementos: 1) la estructura precisa; 2) la actividad individual y las circunstancias (en las cuales la palabra final proviene del entorno físico y humano así como del azar), y 3) las presiones autoorganizadoras que surgen de la misma complejidad del sistema. El perfil impredecible de las experiencias de cada individuo marca el diseño de los circuitos —tanto directa como indirectamente— mediante la reacción que provoca en la circuitería innata y las consecuencias que esas adaptaciones reactivas tienen en el proceso global de configuración de los circuitos.[17]
En el capítulo 2 afirmé que la operación de los circuitos neuronales depende del patrón de conexiones entre las neuronas y de la fuerza de las sinapsis que hacen esas conexiones. En una neurona excitativa, por ejemplo, las sinapsis fuertes facilitan la descarga en tanto que lo opuesto vale para las débiles. Ahora puedo decir que —como diferentes experiencias afectan la relativa fortaleza de las sinapsis en y a través de muchos sistemas neuronales— la experiencia configura el diseño de los circuitos. Por otra parte —en algunos sistemas más que en otros— la fuerza relativa de las sinapsis puede cambiar en el curso de la vida, reflejando diferentes experiencias del organismo; el diseño de los circuitos cerebrales continúa cambiando. Los circuitos no sólo son receptivos a la primera experiencia, sino que conservan plasticidad y son modificables por nuevas experiencias.[18]
Algunos circuitos se remodelan una y otra vez a lo largo de la vida, conforme a los cambios que experimenta un organismo. Otros permanecen generalmente estables y forman la columna vertebral de las nociones que construimos del mundo interno y externo. La idea de que todos los circuitos son evanescentes es absurda. Una plasticidad global habría creado individuos incapaces de reconocerse entre sí y carentes de biografía propia. Eso no sería adaptativo, y evidentemente no sucede. Una prueba sencilla de que algunas representaciones adquiridas son relativamente estables se encuentra en la condición conocida como «miembro fantasma». Algunos individuos a quienes se les amputa una extremidad (por ejemplo, la mano y el brazo, lo que los deja con un muñón arriba del codo), dicen a sus médicos que aún la sienten en su lugar, que pueden percibir sus movimientos imaginarios y sentir dolor, frío o calor «en» el miembro que ya no tienen. Obviamente, estos pacientes poseen memoria de su extremidad perdida: si no fuera así no podrían formar su imagen en la mente. Sin embargo, con el tiempo, el fantasma se atenúa, lo que parece indicar que la memoria —o su reproducción en la consciencia— está siendo revisada.
El cerebro necesita un equilibrio entre circuitos cuya inclinación a la descarga cambie tanto como el mercurio y otros que sean más resistentes si bien no impermeables al cambio. Los circuitos que en este momento nos ayudan a reconocer nuestro rostro en el espejo sin sorpresa han ido cambiando sutilmente para adaptarse a las modificaciones estructurales que el tiempo nos ha grabado en el rostro.