CAPÍTULO
2
La paradoja humana
He utilizado más o menos indistintamente los términos consciencia y conciencia, aunque suelo usar conciencia (por ejemplo en «conciencia visual») para algún aspecto concreto de la consciencia. Hay filósofos que distinguen entre ambos términos, pero no hay acuerdo general sobre cuál es la diferencia. Debo confesar que, en una conversación, suelo decir «consciencia» cuando deseo sobresaltar a mis interlocutores y «conciencia» cuando intento no hacerlo.
Francis Crick, La búsqueda científica del alma
El invento
La adaptación del Homo habilis a los ecosistemas despejados y herbosos, las sabanas, no sólo supone un cambio de hábitat, sino también un cambio de nicho ecológico, es decir, del papel que la especie juega en la cadena de la vida: en otras palabras, de cómo se ganan la vida sus miembros. Por primera vez la carne y las grasas animales pasaron a formar una parte importante de la dieta de los homínidos. Sin embargo, y sorprendentemente, tal mudanza de nicho ecológico no parece suponer una modificación esencial en la morfología del Homo habilis, que permaneció muy semejante a los australopitecos, como hemos visto. A pesar de todo, hay ligeros cambios en la cabeza, con una cara un poco más pequeña y un cerebro algo más grande.
El aumento del tamaño del cerebro puede tener que ver con la nueva forma de vida, basada en la explotación de recursos más dispersos y menos previsibles que los del bosque tropical. Este razonamiento es válido para los productos vegetales y, todavía más, para los animales. El cerebro ampliado otorgaría al Homo habilis una capacidad adicional para almacenar en el interior de la cabeza la cartografía de un territorio muy amplio, es decir, para hacer detallados mapas mentales, y además para interpretar las huellas de los animales y otros signos, como el vuelo de las aves carroñeras cuando han detectado un animal muerto. Acaso también para entender los ritmos de la vida y de la tierra, los cambios de las estaciones entre ellos, para adelantarse a los acontecimientos (previsibles) del mundo natural y para planificar a más largo plazo. Si esto fuera cierto nos encontraríamos ante un cambio realmente muy significativo, porque no parece que los chimpancés hagan ningún tipo de planes para el futuro. Es asimismo muy verosímil que los grupos sociales se hicieran más amplios, más integrados, más cooperativos, y que el incremento de materia gris responda a un aumento de la complejidad social, es decir, que sirva también para adelantarse al comportamiento de los demás.
El primatólogo Robin Dunbar ha estudiado el tamaño del cerebro y de sus partes constituyentes en los primates para ver con qué variables se corresponden los grandes cerebros que se observan en muchas de sus especies. Después de descartar otras hipótesis, Robin Dunbar se quedó solamente con dos: ¿el tamaño del cerebro se relaciona sólo con el nicho ecológico o, más bien, con el tamaño y complejidad del grupo social? El resultado final de sus estudios es que existe una asociación muy estrecha entre la complejidad social de un primate y el tamaño de su neocórtex, y que no existe ninguna variable ecológica que haga aumentar el tamaño del neocórtex. El neocórtex representa en los humanos la mayor parte del cerebro, pero sin embargo no es la parte dominante en los reptiles y en los mamíferos que no son primates.
El aumento del neocórtex del Homo habilis sería, por lo tanto, un fenómeno social. Se puede añadir a esto que hubo otros homínidos, los parántropos (de los que hablaremos luego), que también se adaptaron, y al mismo tiempo que el Homo habilis, a los ecosistemas abiertos, sin que ello implicara un gran aumento de su tamaño cerebral. Sin embargo, dado que el incremento del neocórtex afecta a las funciones mentales de asociación y capacidad de análisis, tengo la certeza de que habría servido a los primeros Homo habilis para desenvolverse tanto en un medio social muy rico como para ocupar un nicho ecológico totalmente nuevo; y es muy posible que su insólita complejidad social fuera la clave de su éxito ecológico, y del nuestro posterior.
Por último, se produce un gran invento. Las innovaciones morfológicas que hemos considerado hasta ahora son producto de la evolución, un resultado del juego entre la mutación y la recombinación —es decir, las fuerzas de la genética—, y de la selección natural —o la fuerza de la ecología si se puede decir así—. Pero ahora la novedad procede de la mente, y por eso puede considerarse el primer invento: la piedra tallada. Los primeros artefactos líticos (o sea, de piedra) datados con seguridad se han recogido en Gona, en la región de Hadar, País de los Afar (Etiopía) y cuentan con unos dos millones y medio de años de antigüedad. Otros conjuntos líticos procedentes del lago Turkana, del río Omo, del Congo, de Uganda y de Malawi parecen tener edades sólo algo inferiores. El primer fósil humano asociado a artefactos comprende la parte inferior de una cara (el maxilar) con el paladar y algunos dientes de hace 2,33 millones de años, encontrado por el equipo de Donald Johanson también en la región de Hadar.
El ambiente que puede deducirse por la abundancia de fósiles de antílopes de pasto era bastante abierto, y claramente menos boscoso que el de los antílopes impala y los Australopithecus afarensis que habitaron antes la misma región. El fósil (A.L. 666-1) es sin duda de un Homo, pero la atribución a la especie Homo habilis de un resto incompleto como éste es sólo una conjetura que me permito. En cualquier caso, estos artefactos de Hadar y otros posteriores asociados a Homo habilis eran cantos toscamente tallados y las lascas desprendidas en la operación (como en realidad no es fácil saber en estos conjuntos primitivos cuáles son los verdaderos instrumentos y cuáles los residuos inservibles de la talla, algunos autores prefieren utilizar el término más amplio de artefactos para abarcar a las lascas y a lo que queda cuando se extraen: el núcleo). Los arqueólogos se refieren a esta industria como Olduvayense, o Modo Técnico I. Se ha dicho que los instrumentos —cualesquiera que fuesen— producidos por el Homo habilis eran «instrumentos biológicos», es decir, que potenciaban o prolongaban la morfología del individuo. Sin embargo, desde que los caninos empezaron a reducirse en el Ardipithecus ramidus, los homínidos no disponen de buenas herramientas naturales para cortar la piel y la carne de los animales muertos, ni tampoco tienen modo de fracturar los huesos para extraer el tuétano, por lo que los instrumentos de piedra realmente representaron la llave, o una de ellas, para abrir una nueva despensa.
Es cierto que los chimpancés son capaces de utilizar instrumentos naturales cuidadosamente seleccionados (y no son los únicos animales que hacen tal cosa), e incluso los modifican ligeramente para adaptarlos a la función deseada. Se los ha observado, por ejemplo, cascando nueces utilizando una roca como martillo y otra como yunque. Pero ningún chimpancé ha sido visto jamás partiendo deliberadamente una piedra, ni en condiciones experimentales se ha conseguido que golpeen con tino una piedra contra otra para producir bordes cortantes. Sin embargo, se les puede hacer comprender la utilidad del filo de las lascas para cortar, y llegan a utilizarlas aunque no sean lo bastante diestros como para confeccionarlas. Todo parece indicar que sus brazos y manos no están bien coordinados para este tipo de actividad (esto no es un grave demérito, ellos son mucho más hábiles que nosotros con los pies). Tampoco los chimpancés tienen mucha puntería a la hora de lanzar objetos, y tallar no es otra cosa que golpear con acierto una piedra contra otra; hace falta buscar el ángulo adecuado y el punto preciso de impacto, y aplicar la fuerza justa. Cuando quieren intimidar a alguien, los chimpancés arrojan palos u otras cosas de una manera bastante torpe, que no tiene nada que ver con nuestra puntería. Por otro lado, es importante señalar que en condiciones naturales no se les presentan a los chimpancés situaciones en las que sea necesario disponer de un filo, por lo que las habilidades (mentales o anatómicas) precisas para crearlos no han sido favorecidas por la selección natural.
Sin embargo, los australopitecos tenían una conformación de brazos y manos esencialmente igual a la nuestra. Seguramente tenían la capacidad mental y biomecánica para confeccionar instrumentos, aunque no se han encontrado en ninguno de sus yacimientos. Es posible que no los necesitasen. Por eso pienso que, pese a lo espectacular de la aparición de los primeros artefactos, no reflejan un salto mental tan trascendental como se ha pensado. Además, esta manera de tallar no parece reflejar la existencia de un modelo ideal de instrumento, que se impone a la piedra como si se tratase de una plantilla mental, sino que simplemente se pretende la obtención, como sea, de un filo: se busca un atributo y no una forma. En todo caso, los filos producidos por los instrumentos fabricados por el Homo habilis les facilitaron la entrada en un nicho ecológico nuevo, el de los comedores de carne. Ésta es la razón de que quizá deba calificar al Homo habilis sólo de «casi humano», ya que su aparición representa en la evolución humana un paso más importante en lo ecológico y en lo social que en lo cognitivo.
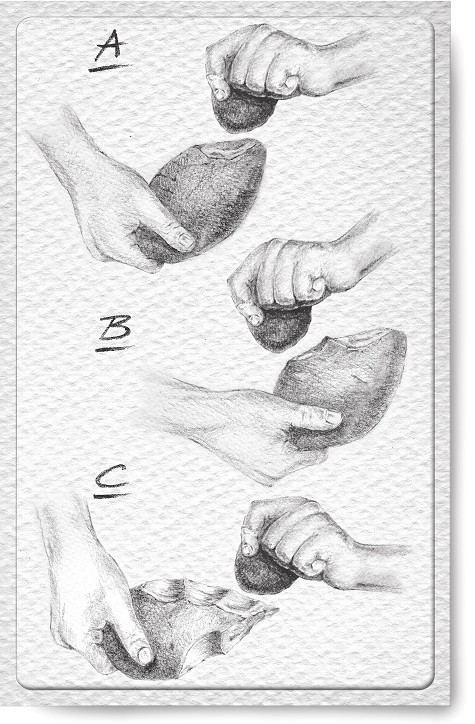
Figura 5: Talla de la piedra. Al golpear un canto con otro una o dos veces por la misma cara (A y B) se obtiene un instrumento simple dotado de filo (chopper); si la talla continúa y se extiende a las dos caras se llega al bifaz (C).
Sin embargo, los primeros fabricantes de utensilios de piedra, hace la friolera dos millones y medio de años, nos han dejado tal vez pruebas de actividad consciente en relación con el uso de herramientas. En algunos yacimientos, la materia prima parece haber sido transportada desde largas distancias (kilómetros) para ser utilizada en la confección de instrumentos, que a continuación se aplicaron al consumo de animales. Aunque nadie ha podido ver tales actividades del remoto pasado, quedan como indicios los instrumentos de piedra abandonados, junto con los residuos de la talla, y las marcas en los huesos de los herbívoros. Cabe pensar que los homínidos iban a buscar las piedras a la vista de un cadáver. Pero, en regiones donde las rocas adecuadas escaseaban, es posible que los homínidos transportaran consigo las piedras o incluso los propios instrumentos, ya preparados, cuando salían a merodear, por si acaso daban con alguna carroña y necesitaban cortar inmediatamente la carne y partir los huesos (y así no correrían el peligro de perderla en la dura pugna que habrían de sostener con los grandes depredadores y con los otros carroñeros).
Ninguno de los animales que utilizan instrumentos, incluidos los chimpancés, exhiben semejantes dosis de previsión, y más bien se preocupan de conseguir un instrumento sólo cuando tienen la necesidad de usarlo. Además, nunca van muy lejos a por él, sino que buscan a su alrededor los materiales, en un radio de pocos metros. Si finalmente se probara que los primeros homínidos no se limitaban a producir un filo, de una manera mecánica, con las piedras que tenían a mano, o a machacar un hueso con un bloque encontrado en las proximidades, sino que mantenían largo tiempo en su cabeza la idea de conseguir la piedra, entonces quedaría demostrada la existencia de comportamiento tecnológico consciente más allá de lo conocido jamás en ninguna especie actual no humana.
Una característica del comportamiento de los animales es que sus acciones van encaminadas a conseguir objetivos inmediatos, que en muchas ocasiones tienen delante de sus ojos. Los chimpancés cazan a veces pequeños monos, sobre todo colobos, entre otros pequeños animales, pero no parece que planeen estrategias para capturar presas que no están a la vista (sino que sólo son hipotéticas); su comportamiento cinegético es más bien oportunista: aprovechan la ocasión cuando se presenta, y más que organizarse en partidas parece que los machos se van sumando a la caza una vez que ha comenzado. También fabrican esponjas vegetales para absorber líquidos, o pelan palitos para introducirlos en los termiteros y capturar insectos, pero siempre con el propósito de utilizar las esponjas o los palitos en seguida.
Se podrá objetar que las aves acarrean durante mucho tiempo los materiales con los que construyen sus nidos, como hacen los castores con sus diques, o las hormigas con sus hormigueros, y así podríamos seguir con todas las clases de construcciones realizadas por animales. Sin embargo, éstos son unos comportamientos totalmente instintivos y por lo tanto preprogramados, y en modo alguno indican perseverancia en la persecución de un objetivo al que se ha llegado como resultado de una toma de decisión. Los panales hechos de celdillas perfectamente hexagonales no indican en modo alguno la existencia de arquitectos entre las abejas. En el caso de los primeros homínidos, si se demostrase que las piedras eran acarreadas desde que se ponían en marcha o, más aún, que los utensilios eran preparados con anticipación, en lugar de fabricados después de hallado el cadáver, aún estaría más firmemente convencido de su capacidad de planificación, es decir, de que los homínidos sabían lo que querían hacer. ¿Cómo podría demostrarse tal cosa? En caso de que se encontrasen muchos huesos de herbívoros con marcas de corte o machacados, y en cambio ninguna piedra tallada en los yacimientos, entonces se podría interpretar tal hecho como que los homínidos apreciaban tanto sus instrumentos, a causa de la escasez de materia prima, que no los abandonaban después de usarlos y se los llevaban consigo. Precisamente, eso es lo que creen haber descubierto Tim White y sus colegas en los mismos yacimientos de Etiopía, datados en 2,5 millones de años de antigüedad, en los que se rescataron los restos de Australopithecus garhi.
El primer humano
Los fósiles más conocidos de Homo ergaster, la siguiente rama del cladograma de la figura 4, son dos cráneos y un esqueleto casi entero procedentes de la cuenca del lago Turkana en Kenia. El cráneo más completo tiene una edad de unos 1,8 millones, y los otros dos fósiles una antigüedad de 1,6 millones de años. Recientemente, un equipo italiano de la Universidad de Florencia ha encontrado en la depresión de Danakil, en Eritrea, un cráneo que, a juzgar por el estudio preliminar, parece de esta misma especie, y que se data en tan sólo un millón de años; la fecha es, como siempre ocurre cuando se acaba de producir un descubrimiento, provisional: habrá que esperar un poco más para estar seguros. De ser cierta, el rango cronológico de la especie Homo ergaster se extendería entre algo menos de dos millones de años y aproximadamente un millón de años. Su distribución geográfica podría haber sido muy amplia, ya que del yacimiento sudafricano de Swartkrans proceden algunos restos fragmentarios que pueden atribuirse a la misma especie; su edad geológica se sitúa en torno al millón y medio de años. En este caso, y por primera vez, nos encontraríamos ante un homínido identificado tanto en el este de África como en el sur del continente.
La mayoría de los fósiles del Homo habilis tienen 1,8-1,9 millones de años, aunque el rango cronológico de la especie se puede extender hasta los 2,33 millones del fósil de Hadar y de otros restos aún menos completos procedentes del río Omo. En principio no hay problemas desde el punto de vista de la cronología ni de la biogeografía para aceptar que esta especie sea la antepasada del Homo ergaster, cuyos fósiles tienen 1,8 millones o son posteriores. Es cierto que algunos especímenes comúnmente considerados de Homo habilis procedentes de la capa II (Bed II) de Olduvai se datan en 1,7-1,6 millones de años y 1,5-1,4 millones de años; los demás fósiles de Homo habilis de la quebrada de Olduvai proceden de la Bed I, la capa más antigua, y tienen unos 1,8 millones de años. También hay un fósil que yo considero de Homo ergaster, un coxal o hueso de la cadera, que podría tener entre 1,8 y 2 millones de años. No obstante, algunos autores creen que los fósiles más modernos de Olduvai (Bed II) están demasiado «evolucionados» para ser de Homo habilis, y acaso sean de Homo ergaster, y por otra parte la procedencia estratigráfica (y la cronología) del coxal del lago Turkana no está del todo clara; tal vez tenga 1,8 millones de años o aún menos.
Hay quienes ven en este posible (que no seguro) solapamiento entre los últimos fósiles de Homo habilis y los primeros de Homo ergaster un problema para aceptar que la segunda especie proceda evolutivamente de la primera. En realidad no tiene por qué coincidir necesariamente la extinción de una especie en todo el mundo con la aparición de su descendiente, también en todo el mundo. Eso sucedería tan sólo si una especie se convirtiera en otra a lo largo de su área completa de distribución geográfica, en un proceso que afectaría a todas las poblaciones sin excepción. En la mayor parte de los casos, empero, la especie descendiente evolucionó en un lugar geográfico concreto a partir de una única población de la especie antecesora, por lo que luego pueden coexistir durante mucho tiempo las dos especies ocupando lugares diferentes (por ejemplo el Homo ergaster en las comarcas del lago Turkana y el Homo habilis en Olduvai). Es más, si la especie descendiente se extiende a otras regiones donde todavía habita la especie antecesora podrían incluso llegar a convivir las dos especies (la madre y la hija). Sin embargo, cuando ambas jueguen el mismo papel en el ecosistema competirán entre sí allí donde convivan y la especie antecesora podría terminar por desaparecer. Este mecanismo, a grandes rasgos, es el que se postula para el tránsito Homo habilis-Homo ergaster y para otras secuencias parecidas en la evolución humana.
El Homo ergaster era muy diferente de todos los homínidos anteriores en muchos sentidos importantes. Para empezar su cuerpo había cambiado, adquiriendo una talla y unas proporciones semejantes a las nuestras, y bien diferentes de las de los australopitecos y primeros Homo. Hay algunos restos aislados que así lo indican, como por ejemplo el hueso de la cadera ya mencionado, o un fémur; ambos fósiles proceden de la margen oriental del lago Turkana. Pero la confirmación de que el patrón corporal moderno surgió en África con el Homo ergaster ha llegado con el hallazgo por parte del equipo de Richard Leakey de un esqueleto asombrosamente bien conservado en la orilla occidental del Turkana (en la localidad de Nariokotome). Perteneció a un muchacho de unos 9-10 años de edad en el momento de morir que tenía una estatura comparable a la de un chico moderno de su edad, si no era aún más alto.
Por otro lado, se produjo un aumento importante del volumen encefálico en Homo ergaster, alcanzando en los cráneos mejor conservados valores de 804 cc, 850 cc y 900 cc. Este crecimiento del cerebro en Homo ergaster, no obstante, se produce al mismo tiempo que un incremento del tamaño y peso corporal, por lo que en términos relativos no representa un avance demasiado grande con respecto al Homo habilis (aunque ya está dicho que es posible que las capacidades craneales de esta última especie se hayan sobreestimado en el pasado).
Sin embargo, me cuesta mucho trabajo aceptar que el incremento cerebral que experimentó el Homo ergaster con respecto al Homo habilis no supusiera también un gran salto en las capacidades cognitivas. Hasta ahora siempre que he dado una cifra de tamaño cerebral de una especie, a renglón seguido he añadido también un comentario sobre el peso del cuerpo. La razón de este proceder es que si una especie aumenta su tamaño corporal cabe esperar que también lo hagan todos los órganos del cuerpo, y el cerebro es uno de ellos, ni más ni menos que el hígado.
Esta estrategia científica funciona bien en términos generales cuando se comparan grupos muy diferentes de mamíferos, con tamaños del cerebro y del cuerpo muy desiguales. A niveles taxonómicos inferiores no es tan útil, en cambio. Para empezar, diferentes personas, o poblaciones humanas, muestran distintos pesos relativos del cerebro, sin que ello exprese diferencias en la inteligencia. En promedio, y a igualdad de peso, el cerebro de un varón es unos 100 cc mayor que el de una mujer, pero que nadie se altere, porque esa diferencia no tiene nada que ver con las funciones mentales llamadas superiores (las capacidades cognitivas). La prueba es que en los macacos, que carecen de la inteligencia humana, también se dan disparidades entre los sexos de la misma proporción. Podría guardar relación con la capacidad de procesar información visual y espacial, que aparentemente es superior en el sexo masculino en nuestra especie, y quizás también entre los macacos. De hecho las diferencias más claras entre varones y mujeres en los tests de los psicólogos se dan en pruebas tales como rotar mentalmente figuras, leer mapas, recordar posiciones de cosas y manejar los conceptos de derecha e izquierda. ¿Tal vez, se preguntan Dean Falk y otros colegas que han hecho esta investigación sobre el dimorfismo sexual en el tamaño encefálico, en los homínidos machos la selección natural favorecía la capacidad de orientación? ¿Seremos los varones en general mejores navegantes? Pero dejemos de especular y volvamos al tema.
El chimpancé común y el gorila presentan cerebros de tamaños mucho más próximos entre sí en términos absolutos que sus pesos corporales. La media del peso cerebral del chimpancé se sitúa sobre los 410 g y la del gorila está en torno a los 500 g. Sin embargo, el peso del cuerpo del chimpancé promedia 33 kg en las hembras y 43 kg en los machos, mientras que los promedios en el gorila son 98 kg y 160 kg, respectivamente. En consecuencia, el peso del cerebro es proporcionalmente mucho menor en el gorila, sin que su inteligencia, por lo que sabemos, sea inferior.
Dicho en otras palabras, dos especies próximas que tienen cerebros de parecido tamaño suelen tener capacidades mentales parecidas (heredadas de un antepasado común), independientemente de que sus cuerpos se hayan hecho más grandes o más pequeños con el tiempo. La razón principal de que esto sea así es que el cerebro es un órgano energéticamente muy caro, muy costoso de mantener: en nuestra especie consume el 20 por ciento de la energía disponible, pese a que no suele pasar del 2 por ciento del peso del cuerpo (diez veces menos). En el chimpancé el cerebro representa el 9 por ciento del gasto energético. En consecuencia, si se produce un aumento del peso del cerebro en el tránsito de una especie a su descendiente es, probablemente, porque era muy importante ese incremento, pese a su enorme precio energético. Si no fuera tan necesario que creciese el cerebro la selección natural habría favorecido el que se mantuviera constante, aunque el cuerpo creciese; es probable que los gorilas se hayan vuelto tan grandes porque se han convertido en folívoros, comedores de hojas y tallos, unas partes vegetales poco alimenticias y que exigen grandes aparatos digestivos para ser procesadas. En sentido contrario, la reducción del peso corporal de una especie no la vuelve más inteligente sólo porque haya conservado su tamaño cerebral.
Además, en relación con algunos aspectos importantes de la fisiología, y a la vez del comportamiento, lo que cuenta es el tamaño absoluto del cerebro, y no el relativo. Me estoy refiriendo a la duración del periodo de desarrollo, que es en nuestra especie el más largo de todos los primates, implicando un prolongado periodo de dependencia y de necesidad de alimentación y cuidados antes de que el individuo llegue a adulto. En ese tiempo también aprende uno a relacionarse con los demás y a vivir en sociedad, recibiendo unas lecciones del máximo valor, porque ningún animal de una especie social, y la nuestra lo es en grado sumo, puede sobrevivir solo.
En todos los homínidos anteriores al Homo ergaster el desarrollo duraba lo mismo que en los chimpancés, o poco más. Desde el punto de vista de la osificación completa del esqueleto el desarrollo se termina entre nosotros sobre los 20 años —aunque el crecimiento en estatura suele acabar algo antes—, mientras que en los chimpancés, gorilas y orangutanes cesa en torno a los 12-13 años. El primer embarazo se produce en las hembras de los chimpancés a los 13 años en promedio, es decir que la madurez sexual coincide a grandes rasgos con el final del desarrollo óseo: cuando éste concluye empieza la vida adulta, y con ella la reproducción. Mientras la hembra de chimpancé era una cría, su peso aumentaba día a día, lo que quiere decir que adquiría con la alimentación más calorías de las que necesitaba para sobrevivir, y con ese superávit de energía iba construyendo su propio cuerpo. Una vez alcanzada la madurez reproductora ya no incrementará su peso, pero con el exceso de calorías hará crecer otra criatura dentro de su cuerpo, como si fuera un órgano más; luego, por un tiempo, durante la lactancia, el hijo crecerá a su costa fuera del útero, hasta que llegue una nueva cría. En cierto modo una hembra siempre está creciendo, primero construyéndose a sí misma, y luego «fabricando» a sus hijos.
En el mundo actual la práctica totalidad de los seres humanos vivimos en sociedades de economía productiva en las que el alimento, las plantas y los animales, no se caza o recolecta, sino que se cultiva o cría. Estas condiciones de vida en cierto modo artificiales, en especial en el Primer Mundo, donde se disfruta durante el crecimiento de una alimentación extraordinariamente rica y variada, pueden haber alterado algunos aspectos de nuestra biología del desarrollo, como el comienzo de la edad reproductora, por lo que es mejor recurrir para las comparaciones a las pocas poblaciones que todavía hoy se alimentan como nuestros antepasados, comiendo animales salvajes y recolectando vegetales silvestres. Por ejemplo, entre los Ache de Paraguay y los Dobe !Kung de Namibia el primer embarazo se produce por lo general hacia los 16 años en el primer caso y los 18 años en el segundo, unas edades muy próximas al final del crecimiento en estatura de las mujeres en las respectivas poblaciones. Volveré a hablar de estos pueblos, de embarazos y de niños más adelante. Ahora nos interesa seguir tratando el tema del cerebro.
Se ha demostrado que existe una relación muy estrecha en el conjunto de los primates entre la duración del ciclo vital, o sea de los diferentes periodos de la vida, y el tamaño del cerebro. Por eso, la infancia, la adolescencia, y la vida total de un chimpancé doblan en duración a las etapas equivalentes de la vida de un macaco, cuyo cerebro es aproximadamente la cuarta parte. Por idéntico motivo nuestro ciclo vital es mucho más largo que el de un chimpancé: nuestra longevidad está ligada a nuestro gran cerebro. Siendo el tamaño del cerebro del Homo ergaster intermedio entre el de los chimpancés y el nuestro, cabe suponer que también fuera intermedia la duración de su infancia, adolescencia y vida total. Y un largo periodo de desarrollo supone también un dilatado tiempo de aprendizaje y preparación para la vida adulta.
A la edad de tres años, tanto un chimpancé y un gorila como un niño humano tienen un cerebro que ya ha sobrepasado las tres cuartas partes del tamaño definitivo. Eso quiere decir que la mayor parte del cerebro crece a expensas de la madre, que proporciona la energía necesaria primero en la gestación y luego durante la lactancia. Lo que queda por crecer del cerebro después del destete es sólo una pequeña parte y por eso tiene sentido preguntarse para qué sirve un desarrollo tan prolongado. La respuesta es que durante ese tiempo se produce la «programación» del cerebro, la instalación de un complejísimo software en un hardware ya construido en sus partes esenciales, si se me permite la analogía con los ordenadores. Por todo lo expuesto concedo mucha importancia a la prolongación de este tramo de la vida, porque la considero un requisito esencial para que fuera posible una sociedad compleja, y también para el desarrollo de una tecnología cada vez más elaborada.
Los bifaces
Hace 1,6 millones de años aparece en África un tipo nuevo de instrumentos de piedra (ahora sí que podemos utilizar el término «instrumento» porque los objetos a los que me voy a referir sin duda fueron confeccionados para ser utilizados como herramientas). Se trata de los bifaces, unos utensilios de gran formato tallados por las dos caras con una perfección y simetría evidentes. Pueden ser de varios tipos: hachas de piedra, hendedores o picos. El ciclo industrial al que pertenecen se llama Achelense o Modo Técnico II y refleja un importante salto tecnológico respecto del Modo I (el Olduvayense), porque esta vez se reconoce la búsqueda deliberada, es decir, consciente, de instrumentos con una forma predeterminada, que sólo existía antes en la mente del autor. Marcel Otte considera un bifaz, a todos los efectos, como una escultura, sin excluir que además de ser funcional (es decir, útil) también responda a un gusto por la belleza (una estética). Estos primitivos seres humanos sabían bien lo que se traían entre manos.
Por su cronología, se piensa que los bifaces fueron fabricados por el Homo ergaster; además, en el yacimiento de hace 1,4 millones de años de Konso, en Etiopía, se encuentran asociados el Achelense y una mandíbula de Homo ergaster. Para entender correctamente la diferencia que existe entre la evolución biológica y los cambios culturales, es importante destacar el hecho de que los primeros fósiles de Homo ergaster pertenecen al ciclo tecnológico del Modo I. Por lo tanto, la invención y extensión del Modo II no representa un cambio biológico hacia una mayor inteligencia. En otras palabras, hace un millón y medio de años algunos Homo ergaster utilizaban una tecnología más elaborada que sus antepasados o que los miembros de otras poblaciones de la misma especie (e incluso que seres humanos posteriores, como veremos). Ahora bien, aunque una nueva tecnología no suponga necesariamente la aparición de una nueva especie, también es cierto que una industria muy compleja no es compatible con una mente muy simple: hay seres humanos que no saben informática, pero es del todo imposible que un mono aprenda a utilizar un ordenador. No es probable, en otras palabras, que el Modo II estuviera al alcance de la mente del Homo habilis.
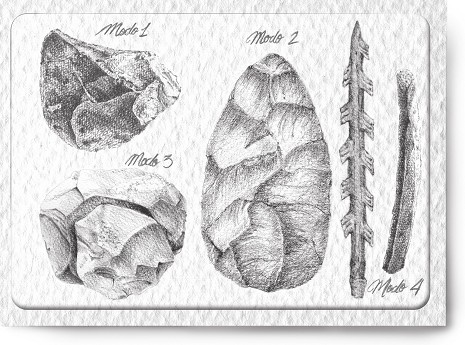
Figura 6: Los cuatro grandes modos técnicos.
El famoso escritor francés Julio Verne publicó en 1901 una novela poco conocida titulada El pueblo aéreo. En ella contaba la aventura de un expedicionario francés y otro norteamericano que encuentran unos seres (los «wagddis»), con características intermedias entre el chimpancé y el hombre, viviendo en un poblado construido en las copas de los árboles, en el dosel de la selva tropical africana. La novela refleja a la perfección el espíritu de los años de exploración del continente africano, cuando continuamente se descubrían tierras con nuevas especies animales y pueblos humanos hasta entonces ignorados en Occidente. Estos hombres-mono que fabuló Julio Verne representaban la supervivencia, en un lugar recóndito e inexplorado de la selva del Congo, de un eslabón intermedio en la evolución humana. En un pasaje de la novela se lee: «En cuanto al microcéfalo, del que se quiere hacer un tipo intermedio entre el hombre y el mono, especie vanamente predicha y buscada por los antropólogos, lazo de unión entre el reino animal y el homal (sic), ¿podía admitirse que estuviera representada por aquellos wagddis?»
Con la especie Homo ergaster nos encontramos también ante unos «eslabones perdidos entre el mono y el hombre» utilizando la jerga del evolucionismo decimonónico, aunque el «contacto» entre nuestra especie y el Homo ergaster se haya producido en un yacimiento y no en una selva remota. Físicamente aquellos humanos fósiles eran muy semejantes a nosotros de cuello para abajo, pero el tamaño de su cerebro era intermedio: cabe imaginárselos con una mente aún muy alejada de la humana moderna, pero también muy alejada de la del primer homínido conocido, el Ardipithecus ramidus, y de los australopitecos (no tanto del Homo habilis). Una mente muy distinta, por buscar un ejemplo próximo, de la del actual chimpancé. Su ciclo vital era más largo en todas sus etapas, aunque todavía no fuera como el nuestro. Eran capaces de fabricar herramientas muy elaboradas que exigen tener una idea clara en la cabeza del resultado que se desea obtener y de los pasos, muchos, que hace falta dar para conseguirlo; no era sólo una cuestión de golpear de cualquier manera una piedra contra otra una o dos veces.
Con el Homo ergaster comienza a manifestarse en toda su tremenda dimensión la extraordinaria paradoja que representan en la biología los seres humanos, las personas: anatómicamente sólo unos primates con postura erguida, una novedad evolutiva interesante pero en ningún sentido más extraordinaria que el vuelo de los murciélagos o la adaptación al mar de los cetáceos; y al mismo tiempo unos organismos radicalmente diferentes de todas las demás criaturas vivientes por su asombrosa inteligencia, capacidad de reflexión y plena conciencia de sí mismos en todos sus actos.
Ecológicamente, el Homo ergaster también era radicalmente distinto de los primeros homínidos: había abandonado definitivamente los ecosistemas densamente arbolados del Ardipithecus ramidus y de los australopitecos, y explotaba los recursos de los medios abiertos. Uno de los máximos especialistas mundiales en los climas del pasado, Peter deMenocal, aprecia en los ecosistemas africanos donde se originó el Homo ergaster una nueva vuelta de tuerca en el proceso de enfriamiento y aumento de la aridez hace precisamente 1,7 millones de años, con expansión de las sabanas y aún mayor reducción del bosque tropical. Además de los vegetales, el Homo ergaster consumía regularmente la carne de otros animales que obtenía en forma de carroña o por medio de la caza. El Homo habilis ya había iniciado este camino, pero su menor corpulencia le haría menos cazador (al menos de presas de medio y gran porte) y más carroñero que el fornido Homo ergaster. Y además, hace menos de dos millones de años sucedió algo muy importante: la evolución humana pasó de ser un fenómeno localizado en un único continente, a convertirse en un suceso de escala mundial; los bípedos pensantes se extendieron más allá de África.
La rama cortada
Pero antes de salir del continente-cuna tengo que referirme brevemente a una rama muy notable del árbol de la evolución humana: la de los parántropos. Éstos eran unos homínidos corporalmente muy parecidos a los australopitecos, hasta el punto de que muchos autores los incluyen dentro de ellos; también son superficialmente similares al Homo habilis, que por lo que sabemos hasta ahora no representaba ninguna novedad en cuanto a la forma del cuerpo. Sin embargo, los parántropos experimentaron una especialización muy notable en su aparato masticador, que se desarrolló extraordinariamente para poder procesar grandes cantidades de productos vegetales duros y fibrosos, muy abrasivos.
Muchos investigadores creen que los parántropos son una respuesta al mismo cambio climático de hace 2,8 millones de años que propició el origen del género Homo, como hemos visto. Esta hipótesis es compatible con la cronología de los parántropos, porque la primera de sus especies, el Paranthropus aethiopicus, se conoce en el este de África hace 2,6 millones de años. Especies de parántropos posteriores son el Paranthropus boisei, también esteafricano, y el Paranthropus robustus, encontrado en algunas cuevas sudafricanas (Swartkrans, Kromdrai y Drimolen).
Una cuestión que no está del todo clara es la de dónde injertar la rama de los parántropos en el árbol de los homínidos. La mayoría de los autores la entroncan a la altura del Australopithecus afarensis, que sería un antepasado común de los parántropos y de los humanos. A efectos prácticos, el auténtico esquema evolutivo puede ser éste o uno muy parecido. Sin embargo, a decir verdad, el Australopithecus afarensis podría no ser exactamente un antepasado común de parántropos y humanos, porque en algunos rasgos nos parece a mi colega Ignacio Martínez y a mí ya «un poco parántropo». En este caso, el antepasado común de las dos ramas sería el Australopithecus anamensis o, como digo siempre, alguna especie muy semejante (a decir verdad, y por lo que sabemos, el Australopithecus afarensis y el Australopithecus anamensis son también dos especies muy parecidas entre sí).
El poblamiento de Asia
Hasta aquí ésta ha sido una historia exclusivamente africana, como la de los gorilas y los chimpancés. El Ardipithecus ramidus, los australopitecos y el Homo habilis nunca conocieron otros paisajes (ni otros animales y plantas) que los que se dan en las cálidas tierras del continente que ha sido nuestro primer hogar. Cada especie animal está adaptada a su nicho ecológico, al lugar que ocupa en el ecosistema al que pertenece. Para ello, la evolución le ha dotado de las herramientas que precisa, y no sólo en su morfología (que es la parte que se «ve» de un organismo), sino también en su fisiología (el funcionamiento de los diferentes sistemas del cuerpo), y en su comportamiento. Hay especies que son poco tolerantes a los cambios de su ambiente; viven siempre en condiciones muy estables, en una especie de burbuja ecológica que constituye todo su mundo. A estas especies les es muy difícil escapar de su hábitat, y por lo general su distribución geográfica llega hasta donde termina éste. Los gorilas y chimpancés no pasan del límite del bosque húmedo donde habitan; ahí está emplazado para ellos el final del mundo.
Otras especies animales son ecológicamente más flexibles, más tolerantes a las variaciones en las condiciones del ambiente, tanto del medio físico (el clima, por ejemplo, o la salinidad en los organismos acuáticos), como del medio biológico (las comunidades o biocenosis en las que se integran). Lógicamente estas especies tienen áreas de distribución geográfica mucho más amplias. Algunas veces la evolución produce una especie que ocupa un nicho ecológico nuevo, bien dentro del mismo ecosistema, bien en otro diferente. Por lo general se trata tan sólo de pequeñas variaciones, que diferencian ligeramente a una especie de sus más próximos parientes. Muy rara vez el cambio ecológico es tan profundo que surge una nueva línea evolutiva que transforma por completo el marco en el que se ha desarrollado hasta entonces su existencia, aunque, desde luego, a veces ocurre: hay mamíferos que se han adaptado a vivir todo el tiempo en el mar, y otros que pasan casi toda su vida en el aire.
Los homínidos que hemos tratado antes del Homo ergaster pertenecen sin excepción al tipo de las especies de hábitat restringido. Unos homínidos, los primeros, vivieron en el bosque, y otros posteriores en la sabana, aunque unos y otros lo hicieran siempre en África. Pero llegar, como ahora veremos, hasta los confines de Asia supone un gran cambio ecológico, y colonizar Europa uno aún mayor, porque ningún otro primate vive con el hombre en este último continente. Sin embargo, para adaptarse a las nuevas y variadas situaciones ecológicas que los humanos encontraron fuera de África no se produjeron modificaciones morfológicas ni fisiológicas espectaculares; la flexibilidad ecológica se la proporcionó un único órgano: su cerebro.
Los primeros fósiles asiáticos se encuentran en Java y China. Aunque Java es actualmente una isla, los humanos llegaron a ella andando. La explicación a esta aparente paradoja reside en que Java (como Sumatra y Borneo) emerge de la plataforma de la Sonda, una gran extensión de fondo submarino de escasa profundidad que durante las glaciaciones, y por efecto del descenso del nivel del mar que producen, estaba unida al continente asiático. En Java son varios los lugares que han proporcionado fósiles humanos. Desgraciadamente no se conoce con claridad, en la mayoría de los casos, la procedencia exacta de los hallazgos, por lo que no es fácil establecer su edad geológica. En los últimos años, un equipo de geocronólogos encabezado por el norteamericano Carl Swisher se esfuerza por construir un marco temporal para el registro fósil de la evolución humana en Java. Los fósiles más antiguos han resultado ser (con todas las precauciones debidas por las razones arriba expuestas): a) un cráneo infantil sin cara (una calvaria) hallado en Modjokerto, que se data (o al menos el terreno del que se supone que procede) en 1,8 millones de años; y b) unos restos craneales incompletos y muy deformados del área de Sangiran, con 1,6 millones de años (aunque de nuevo hay que repetir que lo que se fecha es el sedimento de donde supuestamente proceden los fósiles). Además de los citados, el área de Sangiran, que es una gran cuenca sedimentaria, ha dado una colección importante de cráneos humanos.
Estas fechas tan antiguas de Java coinciden con las de los primeros Homo ergaster de África, y podría tratarse de la misma especie. El problema es que el niño de Modjokerto murió demasiado joven (3 a 5 años) para poder saber qué tipo de humano representaba, y los fósiles de Sangiran datados son escasamente informativos: la verdad es que no sabemos muy bien quiénes eran o cuándo llegaron los primeros humanos a Java.
Otro yacimiento javanés es Trinil. Aquí descubrió en 1891 Eugène Dubois una bóveda craneal junto con una muela y un fémur. A partir de estos restos Dubois creó la especie Pithecanthropus erectus. Hoy se llama a la especie Homo erectus, pero el viejo nombre Pithecanthropus erectus respondía a lo que se creía que la especie representaba: un «hombre-mono», un ser erguido que todavía no es en sus características mentales como nosotros, pero que ya lo es en la forma de su cuerpo. Después de todo, Dubois no andaba muy descaminado.
Las capacidades craneales de todos estos fósiles javaneses no van mucho más allá de las de los fósiles de Homo ergaster y varían entre 813 cc y 1059 cc. Recientes estudios de Carl Swisher indican que todos los fósiles de la cuenca de Sangiran tienen más de un millón de años, cualquiera que sea su procedencia. Los fósiles humanos de Trinil, por la fauna a la que estaban asociados, probablemente se correspondan con los ejemplares más modernos de Sangiran.
Hay muchos autores que consideran que no debe hacerse distinción entre los fósiles de Homo ergaster y los de Homo erectus, y que todos ellos serían Homo erectus (siempre prevalece el nombre más antiguo, y éste es del siglo XIX). Yo mismo he realizado un estudio de la arquitectura del cráneo cerebral y he llegado a la conclusión de que no hay diferencias importantes entre unos y otros. Sin embargo, desde el principio los cráneos asiáticos son más robustos —extremadamente robustos en algunos casos— que los africanos, y hay algunas otras diferencias en la base del cráneo que podrían justificar la distinción entre especies. Ahora bien, quiero dejar bien claro que éstas son discusiones entre especialistas, ya que se trata en los dos casos, fósiles africanos y fósiles asiáticos, esencialmente del mismo tipo de homínido. A los efectos de las cuestiones que se tratan en este libro podría haber llamado tranquilamente Homo erectus a los fósiles de Homo ergaster del lago Turkana.
El poblamiento del continente asiático tuvo que ser sin duda anterior al de Java, porque los humanos hubieron de recorrer medio mundo para llegar hasta la isla. Hay un yacimiento en China, la Cueva del Dragón (Longuppo) que tal vez haya proporcionado utensilios líticos y dos fósiles humanos de hace casi dos millones de años, aunque no está del todo claro. Los supuestos utensilios son dudosos, y uno de los restos humanos (un fragmento de mandíbula con dos dientes) podría ser de un orangután fósil. Puestos a dudar, también podemos preguntarnos si el otro fósil, un incisivo humano sin duda, es tan antiguo como el resto del yacimiento o más moderno.
En Dmanisi (Georgia), al sur del Cáucaso, se ha encontrado una mandíbula humana que tiene una edad comprendida entre 1,5 y 1,2 millones de años. Hay en China un cráneo (Gongwaling) y una mandíbula (Chenjiawo) que parecen estar en torno al millón de años. El cráneo está bastante deformado e incompleto, pero por lo que se adivina se trata de un ejemplar de Homo erectus, la misma especie que poblaba Java en esa misma época. Pero la mayor colección de cráneos (en realidad calvarias) de Homo erectus del continente asiático procede de la cueva de Zhoukoudian, cerca de Beijing (Pekín). Estos fósiles parecen abarcar un dilatado espacio de tiempo, entre 300 000 años y 600 000 años, aunque algunos autores suben un poco este intervalo y otros lo bajan. Franz Weidenreich, el científico que los estudió, pudo en cinco casos estimar con bastante exactitud la capacidad craneal de los fósiles de Zhoukoudian, y encontró que variaba entre 915 cc y 1225 cc.
Beijing se encuentra en una latitud próxima a la de Madrid (ambas capitales se sitúan sobre el paralelo 40° N) y los ocupantes de la cueva vivían en unos ambientes bastante similares a los de sus contemporáneos europeos. En su momento conoceremos estos ecosistemas en detalle, pero se puede avanzar que eran muy diferentes de los medios africanos ancestrales o los bosques tropicales de Java.
Prácticamente todos los fósiles humanos de Zhoukoudian se perdieron durante la Segunda Guerra Mundial, aunque de uno de ellos, el Cráneo V, se han encontrado nuevos restos en excavaciones posteriores. Este fósil es importante, ya que es el más moderno de la colección, y sin embargo la capacidad craneal no parece ser superior a la del resto de la muestra. Otro fósil chino de Homo erectus es la calota de Hexian, cuya capacidad craneal no rebasará mucho los 1000 cc.
En la isla de Java son escasos los artefactos encontrados hasta la fecha. Además, como sucede con los fósiles humanos, su procedencia y cronología es, por lo general, problemática. Una excepción positiva es el yacimiento de Ngebung, en el área de Sangiran, donde un equipo franco-indonesio ha encontrado en los últimos años algunos utensilios en un contexto geológico claro, asociados además a un diente humano. Los utensilios son poco elaborados, del tipo de poliedros o esferas. Sin embargo, hay que tener en cuenta que la isla de Java no ofrece rocas adecuadas para tallar objetos más refinados. No ocurre lo mismo en China, por el contrario: en Zhoukoudian se han encontrados miles de artefactos; como en Java, faltan los bifaces, por lo que se ha pensado que el Homo erectus no fabricaba la industria Achelense, tal vez porque nunca necesitó fabricar bifaces, quizá porque salió de África antes de que allí se originase el Modo Técnico II, y luego permaneció aislado de las poblaciones humanas que sí conocieron este tipo de instrumentos. Una tercera hipótesis sugiere que tenemos que hacer una lectura diferente del registro arqueológico del Extremo Oriente. Eso es lo que proponen el chino Huang Weiwen y el estadounidense Rick Potts: han descubierto utensilios de hace 700 000-800 000 años en el sur de China que, aunque no son propiamente achelenses, ellos consideran ya del Modo Técnico II.
Pero mientras que el Homo erectus se enseñoreaba del Lejano Oriente, otras dos evoluciones diferentes, con distintos protagonistas, tenían lugar en Europa y África. Al final de este libro, casi al final de la prehistoria, los tres actores del reparto se encontrarán en un drama que tiene como escenario todo el Viejo Mundo. El nombre del personaje europeo es el de hombre de Neandertal: en los próximos capítulos intentaremos conocer su pasado, antes de que se produjera El Gran Contacto con nuestros antepasados venidos de África. Tenemos la fortuna de que la Península Ibérica jugó un papel importante en esta historia, y una parte fundamental de la información de que disponemos procede de un yacimiento español: la Sierra de Atapuerca.