CAPÍTULO
1
La especie solitaria
El Hombre, tal como la Ciencia consigue hoy reconstruirlo, es un animal como los demás, tan poco diferenciable, por su Anatomía, de los Antropoides, que las modernas clasificaciones de la Zoología, volviendo al punto de vista de Linneo, la incluyen junto con aquéllos en la misma superfamilia de los Hominoides. Ahora bien: a juzgar por los resultados biológicos de su aparición, ¿no es justamente algo muy diferente?
Pierre Teilhard de Chardin, El fenómeno humano
Tan semejantes, tan diferentes
Nos hemos quedado solos en el mundo. No hay ninguna especie animal que se parezca verdaderamente a la nuestra, ya que somos únicos. Un abismo nos separa en cuerpo y sobre todo en mente del resto de las criaturas vivientes. Ningún otro mamífero es bípedo, ninguno controla y utiliza el fuego, ninguno escribe libros, ninguno viaja por el espacio, ninguno pinta cuadros, y ninguno reza. Y no se trata únicamente de una cuestión de matiz, sino de todo o nada: es decir, que no hay animales que sean medio bípedos, hagan pequeños fuegos, escriban frases cortas, construyan rudimentarias naves espaciales, dibujen un poco o recen de vez en cuando.
Esta absoluta originalidad que se da en nuestra especie no es frecuente en el mundo viviente. Por lo general, toda especie forma parte de un grupo de especies similares. Se observa así una continuidad en la naturaleza de unas especies a otras. Sin embargo, esa continuidad se rompe entre los grandes grupos de organismos. No hay en la actualidad una forma intermedia entre las aves y los reptiles, o entre los reptiles y los mamíferos. Tampoco los anfibios pueden considerarse medio peces y medio reptiles. A cada una de esas formas distintas de vertebrados se les da en las clasificaciones tradicionales la categoría de clase, salvo en el caso de los peces, que en realidad forman tres clases distintas: la de los peces óseos (los «normales»), la de los peces cartilaginosos (como los tiburones y las rayas), y la de las lampreas (una clase muy mermada en la actualidad, pero que fue la primera en aparecer). Los vertebrados representan la mayoría de las especies del gran conjunto de los cordados, al que se asigna la categoría superior de filum, que es la más alta en la jerarquía de los animales según el sistema de clasificación que desde los tiempos de Linneo se emplea en Biología.
A su vez, los cordados son radicalmente diferentes de los distintos tipos de invertebrados, como esponjas, corales, equinodermos (el grupo de los erizos y las estrellas de mar), anélidos (las lombrices, por ejemplo), artrópodos (insectos, crustáceos, arañas), moluscos (bivalvos, caracoles, pulpos), y otros muchos tipos o fila (el plural de filum) menos conocidos de invertebrados. Cada una de estas grandes categorías está aislada morfológicamente de las demás.
La antigua doctrina religiosa del origen divino de las especies no daba una explicación satisfactoria para esta coexistencia en la biosfera de especies que forman racimos, que a su vez pertenecen a grupos más amplios que muestran enormes diferencias de diseño entre sí. ¿Es que acaso Dios era un creador tan escaso de imaginación que sólo fue capaz de inventar un reducido número de modelos, a partir de los cuales se vio obligado a desarrollar variantes?
La teoría de la evolución da una respuesta distinta, y más convincente, a este problema: las especies parecidas descienden de un antepasado común cercano en el tiempo, es decir, están estrechamente emparentadas. Por el contrario, los grandes tipos —fila— de organismos surgieron hace muchísimo tiempo, y tienen antepasados comunes muy remotos. Después de tan largos períodos de evolución independiente, es lógico que no se parezcan.
Los primeros vertebrados fósiles tienen más de 450 millones de años, los primeros anfibios más de 350 millones de años, los primeros reptiles más de 300 millones de años, los primeros mamíferos más de 220 millones de años, y más de 150 millones de años las primeras aves fósiles. Sin embargo, desde la aparición de las aves la evolución no ha producido ninguna novedad realmente espectacular. ¿Será que ha agotado su capacidad de innovación? A decir verdad, no hay ningún método exacto para decidir cuándo a un grupo de especies se le da la jerarquía de filum y cuándo la de clase u otra inferior. Se sobrentiende que un filum es una gran categoría que corresponde a un diseño biológico original y muy diferente de cualquier otra forma de organismo del mismo reino. Evidentemente un nuevo filum puede surgir en cualquier momento de la historia de la evolución, ya que no hay ningún motivo para pensar que las cosas importantes sólo ocurrieron en el pasado muy remoto. La razón por la que los mamíferos no constituyen en Zoología un filum propio, y sólo son una clase, es que existen hoy otros organismos con esqueleto, con los que nos agrupamos en el filum de los cordados. Pero eso no quiere decir que los mamíferos no representen un tipo biológico realmente original. En cierto modo, lo mismo nos ocurre a nosotros los humanos, que por el desarrollo de nuestra inteligencia hemos entrado en una dimensión nueva de la biología. El paleontólogo y filósofo francés Pierre Teilhard de Chardin creía que nos correspondía propiamente la categoría de filum.
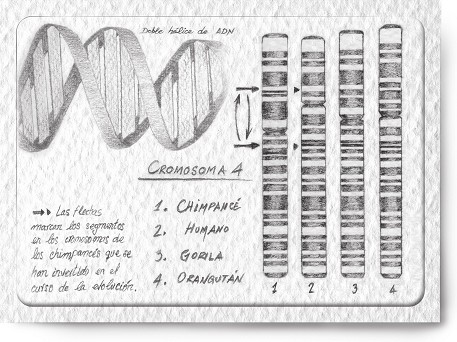
Figura 1: Cromosoma 4. La semejanza genética entre chimpancés comunes, gorilas, orangutanes y humanos no deja lugar a dudas sobre el gran parentesco que existe entre todas estas especies, aunque en el chimpancé se haya producido una inversión en una parte del cromosoma.
Ahora bien, si somos tan diferentes del resto de los mamíferos, ¿quiere eso decir que llevamos mucho tiempo evolucionando por separado? En absoluto. Nuestra estirpe no es, ni muchísimo menos, de las más viejas: no pasa de unos escasos cinco o seis millones de años. Por aquel entonces se separaron las líneas que dieron lugar a los chimpancés por un lado y a nuestra especie por el otro. La escisión de la línea de los gorilas se produjo muy poco antes. ¿Cómo se explica entonces el profundo foso que nos separa de las demás criaturas? La respuesta es doble: por un lado, en algunas características hemos evolucionado muy deprisa, cambiando mucho en poco tiempo; por otro lado, todas las formas intermedias (es decir, de características intermedias) entre nosotros y los chimpancés han desaparecido.
He comenzado este capítulo recordando algunas de las principales diferencias entre el ser humano y el resto de los animales. De entre ellas sólo la postura erguida es un rasgo morfológico, las demás son todas de otra naturaleza, y, en definitiva, se relacionan muy directamente con un único órgano de nuestro cuerpo: el cerebro. ¿Es posible, después de todo, que no seamos tan diferentes de los chimpancés? A decir verdad, sólo nos separa aproximadamente un 1,6 por ciento de nuestros 60 000 a 80 000 genes. Más aún, se estima que no son más de 100, quizás 50, los genes responsables de las diferencias cognitivas entre ellos y nosotros. Un cambio genético pequeño, pero no insignificante sino por el contrario muy significativo, nos convirtió en una especie radicalmente diferente de todas las demás, con propiedades mentales únicas, y no en una mera variante de lo ya conocido y muchas veces repetido. No somos una especie más de chimpancé, sino algo muy distinto. Sin embargo, el zoólogo clasifica a las especies animales por su morfología, y modernamente también por sus genes, así que ¿por qué no nos olvidamos por un momento de nuestra mente y nos comparamos a los demás animales desde el punto de vista de la morfología? Situémonos en la sala de disección del laboratorio y examinemos los cuerpos muertos, sin mente, de las diferentes especies de primates.
Un cuerpo sin mente
El esquema superior de la figura 2 muestra quiénes son nuestros más próximos parientes. El más cercano es el chimpancé, o mejor, las dos especies de chimpancé que existen. Un poco más alejado está el gorila, y algo más el orangután. Los pequeños gibones son nuestros parientes más lejanos dentro de este grupo, como fácilmente comprenderá cualquiera que los vea en un zoológico. Chimpancés, gorilas y orangutanes tienen un aire común, y clásicamente se los agrupaba en una misma familia, la de los póngidos. A los gibones se los incluía a veces entre los póngidos, aunque algunos autores los situaban en su propia familia, la de los hilobátidos. Finalmente, la especie humana era la única de su familia, la de los homínidos. En inglés hay un término común para todos los póngidos (gibones incluidos), y es el de apes, que podemos considerar equivalente al español de «antropomorfos». Homínidos y antropomorfos nos clasificamos todos juntos en la superfamilia de los hominoideos.
Este esquema, que muestra las relaciones evolutivas de un conjunto de primates, es un dendrograma, o diagrama de árbol. Como todas las especies son actuales, y no hay ninguna fósil, el árbol no se puede considerar una genealogía de especies o filograma: faltan los nombres de todos los ancestros, aunque los antepasados comunes a dos o más especies actuales están representados por los puntos de ramificación (nodos o entronques: A, B, C, D, E). En el dendrograma, las especies se unen entre sí en una secuencia determinada, que representa el orden en el que se han ido produciendo en el tiempo las sucesivas separaciones de las líneas evolutivas. Cuanto más alto está un punto de ramificación, más reciente es. En este caso, la más moderna de todas las divisiones es la escisión de los chimpancés en dos especies (E), que viven desde hace unos dos millones y medio de años separadas por el río Congo. El dendrograma no contiene más información, y puede dibujarse de muchas maneras diferentes sin que cambie nada.
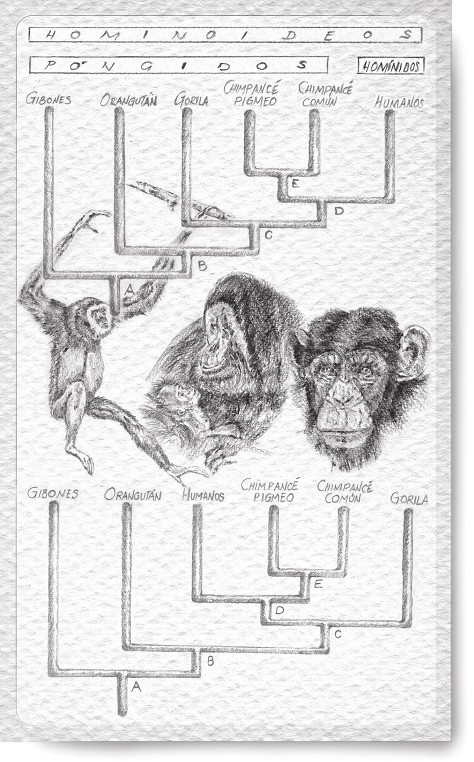
Figura 2: Dos cladogramas equivalentes de los hominoideos actuales.
El dendrograma inferior de la figura 2 es en realidad el mismo que el superior, aunque la posición en que aparece la especie humana haya cambiado notablemente. Ya no está en un extremo, al margen de los demás hominoideos, sino entre ellos. Se comprende ahora que la separación de póngidos y homínidos era artificial, porque en realidad chimpancés y gorilas están más próximos a nosotros, en cuanto a parentesco, que a los orangutanes y gibones. O, en otras palabras, humanos, chimpancés y gorilas tenemos un antepasado común, una especie de «abuelo» (C), del que no descienden los orangutanes y los gibones. Además, el antepasado común de todos los póngidos, el hipotético A, fundador de la dinastía, también era nuestro ancestro. Si fuéramos consecuentes, tendríamos que llamarnos a nosotros mismos póngidos. La única alternativa posible es considerar humanos a los póngidos. ¿Habría entonces que concederles también «derechos humanos»?
Todo lo dicho llama mucho la atención porque pone de manifiesto que parentesco evolutivo y semejanza morfológica no necesariamente son la misma cosa: el chimpancé está evolutivamente más cerca del hombre y, sin embargo, se parece superficialmente más al gorila, y también al orangután. Fue sobre todo el entomólogo alemán Willi Hennig quien se dio cuenta de que para establecer las relaciones evolutivas entre las especies no podemos basarnos exclusivamente en el parecido, y tenemos que hilar más fino. Este hallazgo, en apariencia tan simple, es verdaderamente genial, porque contradice la aparente lógica de que las especies más semejantes deben agruparse siempre juntas. Sólo los grandes sabios son capaces de mirar la otra cara del espejo, y descubrir lo que los demás mortales no podemos ver aunque esté delante de nuestras propias narices.
Un caso similar al de los humanos, aunque a una escala mucho mayor, se da con las aves. Éste es un grupo, una clase, con numerosas especies, cuyos más próximos parientes eran los dinosaurios, concretamente unos pequeños dinosaurios bípedos y carnívoros dentro del grupo de los terópodos (grupo al que también pertenecían otros dinosaurios mucho más grandes y conocidos, como los tiranosaurios). Mejor sería, por lo tanto, decir que las aves son dinosaurios vivientes, los únicos que quedan; las hay grandes como los avestruces —y los humanos hemos conocido aves aún mayores— y pequeñas como los colibríes. Ni siquiera son las aves los únicos dinosaurios con plumas, sino uno de los grupos de dinosaurios terópodos con plumas. Es probable que los famosos Velociraptor de la película Parque Jurásico también estuvieran emplumados en lugar de tener el cuerpo cubierto por escamas; y es posible que, como las aves, también fueran endotermos, es decir, de «sangre caliente»: las plumas parecen haber surgido como una adaptación que ayuda a mantener constante la temperatura corporal, ya que forman un magnífico aislante térmico. La situación actual de las aves es similar a la que se daría si desapareciéramos todos los mamíferos menos los murciélagos. Un hipotético observador del resultado de esa catástrofre sólo podría contemplar mamíferos voladores.
Las aves están aisladas de los demás vertebrados desde que hace 65 millones de años una gran hecatombe acabó con todos los dinosaurios o, si se prefiere, con los dinosaurios que no eran aves. Sin embargo, nuestro aislamiento es mucho más reciente, porque todavía «éramos» antropomorfos hace 7 millones de años; en realidad aún no «éramos» nada, porque por aquel entonces no se habían producido las dos ramificaciones sucesivas que separaron las líneas de los gorilas, los chimpancés y los humanos.
Pero si el estudio de los cuerpos sin mente nos enseña cuál es nuestro lugar entre los primates, también nos muestra que somos diferentes de nuestros más próximos parientes, los chimpancés, porque nosotros somos bípedos y ellos cuadrúpedos, y todo nuestro cuerpo y nuestro esqueleto reflejan esa diferencia en la locomoción.
Pongamos ahora los fósiles, para tratar de llenar el espacio morfológico que nos separa de los chimpancés.
Hombres-mono
En el dendrograma de la figura 3 los nombres comunes de las especies han sido sustituidos por sus nombres científicos en latín: Pan paniscus y Pan troglodytes en el caso de los chimpancés, Gorilla gorilla en el del gorila, y Homo sapiens en el nuestro.
Entre los chimpancés y el hombre aparecen ahora cuatro nuevas especies: Ardipithecus ramidus, Australopithecus anamensis, Australopithecus afarensis y Australopithecus africanus. Ninguna de ellas existe hoy en día, porque desaparecieron hace más de dos millones de años. Las cuatro son especies de homínidos, es decir, que pertenecen a nuestra propia línea evolutiva o estirpe, ya que son posteriores a la ramificación que separó nuestro destino evolutivo del de los chimpancés.
Obsérvese que en el dendrograma no aparece ninguna especie fósil de chimpancé. La razón es que no se conoce ninguna. Sin embargo, no cabe esperar que los chimpancés fósiles vengan a rellenar el foso que nos separa de sus descendientes vivos, por lo que no son importantes en esta discusión: nadie cree que haya habido en el pasado chimpancés más bípedos o más inteligentes que los actuales. Lo que se necesita son formas de algún modo intermedias, «eslabones perdidos» en la retórica tradicional, o dicho aún más crudamente: «hombres-mono».
Éste de la figura 3 es un esquema que contiene tanto especies fósiles como vivientes, pero donde todas aparecen arriba a la misma altura, por lo que no se trata de un filograma o genealogía de especies. Ninguna especie aparece como antepasada de otra. Lo que expresa el dendrograma son los diferentes grados de relación evolutiva o parentesco entre las diferentes especies. Intencionadamente los homínidos fósiles se han situado entre los chimpancés y el ser humano. Ya hemos visto antes que la posición de derecha a izquierda de las ramas del dendrograma es irrelevante, y que lo que importa es cómo se conectan por abajo entre sí las especies, por lo que la posición intermedia de los homínidos fósiles es puramente arbitraria desde el punto de vista de las relaciones filogenéticas (es decir, evolutivas). Sin embargo, desde la perspectiva morfológica las cuatro especies de homínidos fósiles ocupan realmente el lugar de los largo tiempo buscados «eslabones perdidos», aunque estos eslabones no sobreviven en ninguna remota selva, sino que se perdieron en el tiempo, un lugar al que es mucho más difícil ir a buscarlos.
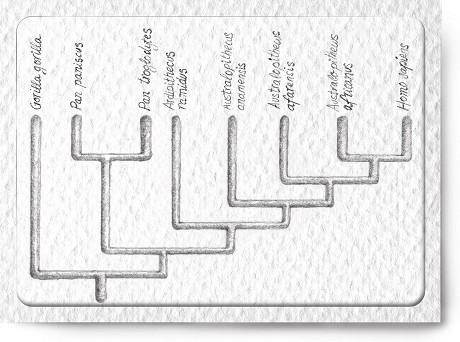
Figura 3: Cladograma que incluye a los australopitecos.
Por decirlo en pocas palabras, la única característica humana que estos homínidos fósiles presentaban (y quizás no todos) era la postura erguida, el bipedismo o bipedalismo, es decir, la unánimemente considerada menos «noble» de nuestras señas de identidad. Su mente era aún la de un mono, la de un antropomorfo como los actuales chimpancés. Eran pues homínidos en el sentido de que pertenecían a nuestra misma familia zoológica, pero no eran aún humanos.
El Ardipithecus ramidus vivió en lo que hoy es Etiopía hace casi cuatro millones y medio de años. El equipo del paleoantropólogo Tim White ha descubierto en los últimos años numerosos fósiles de esta especie, que están en su mayoría aún investigándose. Todavía no se ha dado a conocer el resultado del estudio del esqueleto del cuerpo (o esqueleto poscraneal), por lo que todo son especulaciones acerca de su tipo de locomoción. Se trata sin duda de un homínido muy primitivo, que vivía en la selva lluviosa o pluvisilva, como los gorilas y chimpancés actuales. Por el tipo de dentición se ha deducido que se alimentaba de los mismos vegetales que los chimpancés, es decir, frutas, brotes, tallos tiernos y hojas frescas. Sabemos por lo tanto que pasaban mucho tiempo subidos a los árboles, para alimentarse y para dormir, aunque se desconoce por el momento cómo se desplazaban de un árbol a otro. El Ardipithecus ramidus presentaba, sin embargo, una característica que lo relaciona con los humanos, y no con los chimpancés: los caninos se habían empezado a reducir. Por este único rasgo se sitúa a la especie Ardipithecus ramidus entre los homínidos.
Al lector puede resultarle demasiado confuso que utilice dendrogramas para explicar la evolución humana, en lugar del clásico árbol de especies o filograma. Me apresuro a anunciar que en un próximo capítulo encontrará el lector uno de esos filogramas, el que más me gusta a mí, pero es mejor que no pase todavía las páginas para llegar hasta él. En realidad, los dendrogramas que aquí se presentan son de un tipo especial, y se llaman cladogramas: para su construcción se siguen los principios de la escuela sistemática de Willi Hennig, llamada comúnmente cladística porque reúne las especies en grupos naturales o clados.
Según esta escuela es imposible estar seguros de que una especie fósil es antepasada de otra, fósil o viviente, ya que nadie es verdaderamente capaz de viajar al pasado para seguir el curso de la evolución. Lo único que puede establecerse científicamente es el grado de parentesco entre las especies, tal como aparece reflejado en el cladograma. Para los cladistas los árboles evolutivos son una pura especulación, carente de rigor científico. No debe entenderse lo anterior como que los cladistas niegan la evolución, sino todo lo contrario. Se consideran a sí mismos los más fieles seguidores de las ideas de Darwin. Simplemente, renuncian a ir más allá de la construcción de cladogramas. A la hora de confeccionar estos últimos sólo se utiliza información procedente de la morfología, y por lo tanto da igual que una especie sea fósil o viviente, y proceda de África o de Australia.
Yo no soy un fanático cladista, y creo que la antigüedad de los fósiles y su origen geográfico son datos científicos muy a tener en cuenta en nuestros análisis de la evolución humana. Cuando se suman a un cladograma permiten construir lo que se denomina un «escenario evolutivo», que consiste en un relato o narración de la evolución de un grupo en el que intervienen todas las piezas de información de que se dispone, tal como yo trataré de hacer en este libro. En nuestro caso, el registro arqueológico también interviene, y mucho, en la elaboración del relato, junto con lo que sabemos del clima y de los demás elementos de los ecosistemas en los que los homínidos han vivido, y también sobre ellos mismos. Aunque la veracidad de un escenario evolutivo como tal no puede ser comprobada, los diferentes elementos en que se basa sí que pueden ser confirmados o desmentidos con nuevas pruebas. Si los cambios que se van produciendo con los sucesivos descubrimientos son sustanciales y numerosos, habrá que escribir otro libro distinto. Pero ya tendré tiempo de plantearme esa posibilidad dentro de unos años; quizá no muchos al ritmo al que avanza la paleoantropología. Hoy por hoy, podemos afirmar que el Ardipithecus ramidus era un homínido muy antiguo y primitivo del este de África, y que nosotros procedemos de esta especie, o de una especie muy similar, que vivió hace unos cuatro millones y medio en África, probablemente en su región oriental. De las características de la especie sabremos mucho más en poco tiempo: será menester tener un poco de paciencia.
El siguiente homínido fósil es el Australopithecus anamensis. De él tenemos un puñado de fósiles que vienen de Kenia, y fueron encontrados por el equipo de Meave Leakey en la cuenca del lago Turkana, en las localidades de Kanapoi (orilla occidental) y de Allia Bay (orilla oriental). Todos los fósiles de Kanapoi (menos una mandíbula) han sido datados recientemente con mucha precisión entre hace 4,17 y 4,07 millones de años. Esta especie presenta unas muelas más grandes y con esmalte más grueso que las del Ardipithecus ramidus, lo que nos dice que además de frutos tiernos también ingerían algunos alimentos vegetales que necesitaban una prolongada masticación y producían un intenso desgaste de las coronas dentales. Estos productos vegetales duros y abrasivos eran probablemente granos y frutos secos. Además, se piensa que aprovecharían igualmente órganos subterráneos de almacenamiento de las plantas, como bulbos, tubérculos, raíces engrosadas y rizomas; las partículas minerales que se meterían en la boca con estas partes vegetales enterradas harían rechinar sus dientes y contribuirían a su desgaste. Todos estos productos vegetales se encuentran en bosques más secos que las pluvisilvas de sus presuntos antepasados de la especie Ardipithecus ramidus, por lo que se piensa que el Australopithecus anamensis había cambiado de hábitat, o mejor, que su hábitat había cambiado, haciéndose más seco. Se ha recuperado, además, una tibia bastante completa en Kanapoi (sólo se ha perdido el tercio central), a partir de la cual se ha llegado a la conclusión de que estos homínidos ya eran bípedos. A falta de otra especie de la misma época, podemos afirmar provisionalmente que se trata de antepasados nuestros. En todo caso, nosotros venimos de unos homínidos del mismo tipo que el Australopithecus anamensis.
Sin embargo, los primeros fósiles de esta especie sólo son doscientos mil años más modernos que los fósiles del Ardipithecus ramidus, lo que plantea una interesante pregunta: ¿es suficiente ese tiempo para que se produzcan los importantes cambios anatómicos y ecológicos que van desde el Ardipithecus ramidus hasta el Australopithecus anamensis? Quizá sí y quizá no, porque la evolución no avanza a un ritmo constante, y a veces camina muy rápidamente y otras veces parece estancarse. Lo que es seguro, no obstante, es que si se encuentran en el futuro restos de Australopithecus anamensis con 4,4 millones de años de antigüedad, el Ardipithecus ramidus dejará de ser considerado nuestro antepasado y será visto como una rama lateral de la evolución humana que no llegó a ninguna parte.
Entre hace unos 4 y 2,9 millones de años vivió otra especie de homínido conocida como Australopithecus afarensis. Sus restos han sido encontrados en Tanzania y sobre todo en Etiopía, en el País de los Afar; el protagonista principal de estos hallazgos es Donald Johanson. Como de esta especie se tiene un registro más amplio que de las anteriores, es perfecta para conocer a estos homínidos primitivos. Sus características dentarias nos siguen remitiendo a una alimentación casi totalmente vegetariana en un bosque seco y con claros. Su postura era erguida, aunque los brazos eran largos en relación con las piernas: todavía podían trepar muy bien a los árboles.
El Australopithecus afarensis era una especie de pequeño tamaño en comparación con el nuestro, y apenas más grande que los chimpancés. Los machos medirían unos 135 cm, o poco más, y pesarían alrededor de 45 kg, y la talla promedio de las hembras se situaría en los 105 cm y el peso en unos 30 kg o algo menos. A tenor de estas estimaciones, la diferencia entre los dos sexos era más acusada que entre nosotros o que en los chimpancés, y más próxima a la de los gorilas: en estos australopitecos el peso de los machos era una vez y media (1,5 veces) el de las hembras, en los gorilas es 1,6 veces, 1,3 veces en el chimpancé común y 1,2 veces en la especie humana.
Los dos fósiles estrellas de la colección de Australopithecus afarensis son un esqueleto femenino muy completo apodado Lucy y un cráneo masculino casi entero. El volumen del cerebro de este cráneo se ha estimado en algo más de 500 cc. Otro cráneo más incompleto parece tener una capacidad claramente por debajo de 400 cc. Se trata, aparentemente, de una especie de homínido con un cerebro escasamente más voluminoso que el de los chimpancés, que tienen un promedio cercano a los 400 cc. Como el peso del cuerpo es semejante, tampoco en términos relativos se puede atribuir al Australopithecus afarensis un cerebro mucho mayor, ni una mente con capacidades superiores a las de un chimpancé. El tamaño del cerebro humano varía entre las personas y entre las diversas poblaciones, porque siendo como es, en definitiva, un órgano del cuerpo, depende en parte del tamaño de éste. Por lo general suele darse como promedio para nuestra especie la cifra de 1350 cc, aunque hay tantos millones de seres humanos y tantas poblaciones distintas que esta cifra es más que nada una convención. En todo caso, interesa saber que el promedio humano femenino no llega a los 1300 cc y el masculino supera los 1400 cc. Eso no significa que los varones sean más inteligentes que las mujeres, como discutiremos en su momento. Por otro lado, aproximadamente el 10 por ciento de los seres humanos actuales bajan de los 1100 cc o sobrepasan los 1600 cc, y son completamente normales.
Una vez más, y como ocurre con las especies anteriores de homínidos, no es posible estar seguros de que el Australopithecus afarensis sea nuestro antepasado directo. Algunos autores así lo piensan, pero otros no, como luego veremos. Sin embargo, esta aparente confusión que envuelve siempre la evolución humana no es tan grave como pudiera parecer. En primer lugar, porque quien quiera verdades absolutas, dogmas incuestionables e inamovibles, debe mirar hacia otro lado, que no es el de la ciencia. Ésta sólo elabora hipótesis, vacilantes aproximaciones a la verdad, que siempre pueden ser modificadas total o parcialmente por la fuerza de los hechos: pero es lo mejor que el espíritu humano es capaz de crear. En segundo lugar, porque excepto para la vanidad de sus descubridores no es tan importante que el Australopithecus afarensis se sitúe en línea evolutiva directa con nosotros: podemos estar razonablemente seguros de que tenemos un antepasado en lo esencial como el Australopithecus afarensis, que habitó en África hace entre 3 y 4 millones de años. Y eso es lo realmente importante. Por cierto que recientemente el paleontólogo francés Michel Brunet ha encontrado restos de Australopithecus de esta misma edad en el Chad, en pleno centro de África, lo que me ha llevado a ser prudente y escribir más arriba que nuestro antepasado vivió en África, y no necesariamente en el este de África como habría escrito hace algunos años.
La siguiente rama del cladograma en dirección hacia la nuestra corresponde al Australopithecus africanus. Sus fósiles se datan entre hace 3 millones y algo menos de 2,5 millones, y esta vez no proceden del este de África, sino de tres cuevas de Sudáfrica: Taung, Sterkfontein y Makapansgat. Físicamente eran similares a los Australopithecus afarensis, y su cerebro no parece haber crecido, al menos apreciablemente. Los tres cráneos mejor conservados, todos del Miembro 4 de Sterkfontein, poseen las siguientes capacidades: 375 cc, 485 cc y 515 cc. El último cráneo da la impresión de tener un gran cerebro y se ha llegado a afirmar que su capacidad craneal excedería los 600 cc. Sin embargo, el ejemplar está deformado por la presión del sedimento, como la mayoría de los fósiles de las cuevas sudafricanas, y se hace necesaria una reconstrucción para corregir la deformación del fósil. Glenn Conroy y otros colegas han utilizado una técnica moderna de la radiografía médica cada vez más empleada con los fósiles humanos. Se trata de la tomografía computarizada (TAC), que permite hacer múltiples radiografías seriadas muy próximas entre sí, como si se cortara el fósil en delgadas lonchas. A continuación estas imágenes bidimensionales se introducen en el ordenador y con los adecuados programas se realiza una restitución tridimensional del objeto, que en la pantalla del ordenador se puede modificar para, en este caso, corregir la deformación. Luego es posible tomar medidas como el volumen cerebral. De este modo se ha llegado a una cifra entre 500 cc y 530 cc, que, a pesar de todo, algunos autores juzgan prudente considerar un valor mínimo.
En octubre de 1998, el veterano paleoantropólogo Phillip Tobias, antiguo director de las excavaciones de Sterkfontein, y su colaborador de toda la vida Ron Clarke, actual director de las mismas, han dado a conocer el descubrimiento de un esqueleto muy completo procedente de un nivel profundo de este yacimiento (el Miembro 2); el esqueleto en cuestión podría ser tan antiguo como Lucy —que tiene 3,2 millones de años— o incluso más viejo, y se remontaría hasta los 3,5 millones de años. La circunstancias que precedieron al anuncio del descubrimiento son rocambolescas, como sucede en ocasiones en el mundo de la paleoantropología. En septiembre de 1994 Ron Clarke identificó, entre fósiles de animales que habían sido extraídos del yacimiento dos años antes, algunos elementos del pie izquierdo de este mismo esqueleto, que fue bautizado entonces como Little Foot («Pie Pequeño»). Clarke y Tobias vieron en él rasgos muy primitivos, compartidos con los chimpancés y propios de un animal al menos parcialmente arborícola, cosa que otros discuten. En mayo de 1997 Ron Clarke encontró más restos del mismo pie en el laboratorio, y entre ellos estaban también los extremos inferiores de la tibia y peroné izquierdos, junto con el extremo inferior de la tibia derecha y un hueso del pie del mismo lado, asimismo pertenecientes al individuo Little Foot. A continuación, en junio de 1997, los ayudantes de Clarke en el yacimiento recibieron instrucciones para intentar una misión imposible: la de buscar en las paredes de la grande, profunda y oscura cueva, un hueso cortado que encajara con el fragmento recuperado de la tibia derecha de Little Foot (algo así como encontrar una aguja en un pajar). ¡Y al cabo de sólo dos días lo consiguieron! Todavía permanece Little Foot en gran parte incrustado en la roca, ya que en la cueva de Sterkfontein los fósiles y la matriz endurecida forman un solo bloque, pero ya se ve que el cráneo está entero. Habrá que aguardar por lo tanto un tiempo para conocer de qué homínido se trata y para confirmar su gran antigüedad y sus presuntas características arborícolas. Va a ser una espera emocionante, porque si el esqueleto de verdad es contemporáneo del Australopithecus afarensis, y de una especie distinta (tal vez una forma antigua de Australopithecus africanus), podría competir seriamente por el puesto de antecesor de todos los homínidos posteriores (es decir, con menos de 3 millones de años); entre ellos nos encontramos nosotros.
De lo que se come se cría
Aunque se cree que el Australopithecus africanus vivía en un ambiente forestal no muy diferente al del Australopithecus afarensis, tenía las muelas más grandes, lo que indica que su alimentación requería una más prolongada masticación: aparentemente se basaba en productos vegetales aún más duros que los que ingerían sus antepasados. Pero, ¿hay alguna forma de conocer directamente la alimentación de los homínidos fósiles?
Cualquiera de los elementos químicos puede presentarse en diversas formas, llamadas isótopos. Por ejemplo, nosotros tenemos en nuestros huesos carbono del tipo C12 y, en mucha menor proporción, también del tipo C13, llamado carbono pesado. La diferencia entre ambos es que el carbono pesado tiene trece neutrones y protones en el núcleo y el carbono ligero doce.
Matt Sponheimer y Julia Lee-Thorp han llevado a cabo un magnífico estudio de las proporciones entre el carbono pesado y el carbono ligero en la comunidad de fósiles del yacimiento de Makapansgat, que se data en unos 3 millones de años e incluye representantes del homínido Australopithecus africanus. En África, los árboles y arbustos retienen menos carbono pesado que las hierbas de los pastizales abiertos. Por eso los ungulados que consumen las hierbas acumulan proporcionalmente más carbono pesado que los que comen las hojas de los árboles. Analizando el esmalte de los dientes fósiles, estos autores han encontrado que —como era de esperar— los australopitecos tenían menos carbono pesado que los animales de pasto como los antílopes redunca y los Hipparion (unos équidos con tres dedos en las extremidades, en lugar de uno como los caballos); sin embargo, los australopitecos tenían más carbono pesado que los habitantes del bosque, como los antílopes de cuernos en espiral del tipo del kudú y del sitatunga. Es posible que los australopitecos de Makapansgat consumieran, además de los frutos carnosos y las hojas tiernas de los árboles, también las raíces y las semillas de las altas hierbas de la sabana. O tal vez se alimentasen de insectos comedores de estas hierbas o de los animales que pastaban en campo abierto: es posible que mataran recentales o buscaran carroña. El crecimiento del tamaño de los molares que se detecta desde el Australopithecus afarensis al Australopithecus africanus me hace pensar más en granos, nueces y órganos subterráneos de reserva que en productos animales; para procesar estos últimos no se necesitaría aumentar la superficie de masticación, sino más bien instrumentos que cortaran la carne y machacaran los huesos, que nunca se han encontrado asociados a estos australopitecos. En todo caso, parece deducirse de lo que nos cuentan los isótopos estables del carbono que los australopitecos de Makapansgat no vivían encerrados en el bosque denso y frecuentaban también ambientes más despejados.
Casi humanos
En el cladograma de la figura 4 he injertado dos nuevas ramas entre los australopitecos y nosotros. Ambas representan a miembros de nuestro mismo género: son, como el hombre actual, Homo. La más alejada de nosotros, la más próxima a los australopitecos, corresponde a una especie llamada Homo habilis, la primera del género. Su registro se extiende por Etiopía (valle del río Omo y Hadar), Kenia (lago Turkana) y Tanzania (quebrada de Olduvai), y va desde hace 2,3 millones de años hasta hace 1,5 millones de años. Es interesante atender a la distribución geográfica de las tres últimas especies de homínidos que hemos tratado, porque si quisiéramos hacer descender al Homo habilis del Australopithecus africanus, y a éste a su vez del Australopithecus afarensis, tendríamos que viajar del este de África (Australopithecus afarensis) a Sudáfrica (Australopithecus africanus), y volver luego al este de África (Homo habilis). La irrupción de la biogeografía en nuestro escenario evolutivo nos ha causado un grave problema.
Algunos autores reconocen fósiles de la especie Homo habilis en las colecciones de Sudáfrica: podría haber evolucionado allí mismo a partir del Australopithecus africanus y extenderse luego al este de África, pero yo no acabo de ver esos supuestos Homo habilis sudafricanos. Además, los primeros fósiles de Homo habilis vienen de Etiopía. Mi hipótesis preferida es que en el este de África existió entre hace 3 y 2,5 millones de años una especie próxima al Australopithecus africanus, a partir de la cual evolucionó el Homo habilis. Después de haberle puesto el punto final al libro, he tenido que volver a esta página porque el equipo de Tim White ha nombrado una nueva especie de homínido, llamado Australopithecus garhi, a partir de restos craneales y dentales que han sido hallados en Etiopía (dentro del territorio Afar, en el curso medio del río Awash y no lejos de las localidades en las que este mismo equipo descubrió los fósiles del Ardipithecus ramidus). Estos restos se datan en unos 2,5 millones de años y parecen encajar bien en el modelo evolutivo en el que yo creía, por lo que es razonable pensar que los primeros Homo habilis proceden de una especie de homínido del tipo del Australopithecus africanus, y que su origen tuvo lugar en África, sin mayores precisiones geográficas: aparte del Chad, el este de África y Sudáfrica, otro posible escenario es Malawi, entre las dos últimas regiones. Tim Bromage y Friedemann Schrenk encontraron en la orilla del lago Malawi una mandíbula de aproximadamente 2,5 millones de años de antigüedad que ellos asignan al género Homo, aunque tampoco tengo por segura esta atribución.
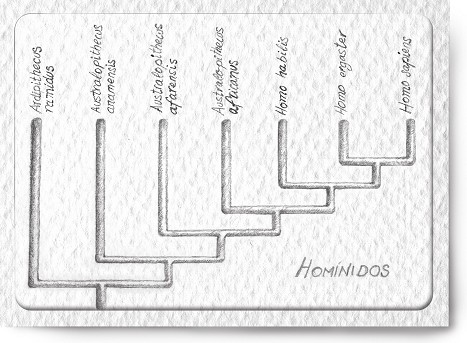
Figura 4: Cladograma de los homínidos. Para simplificar, los parántropos no se han incluido en el cladograma.
El cuerpo del Homo habilis no era diferente del de los australopitecos: pequeño de estatura, los brazos eran largos y las piernas cortas. Al menos eso es lo que parece indicar el esqueleto más completo de que se dispone, procedente de Olduvai (Tanzania) y encontrado por Donald Johanson y Tim White. Otro esqueleto parcial, éste descubierto por el equipo de Richard Leakey en el lago Turkana, es similar. Desde el punto de vista de la morfología no hay muchos motivos para aceptar al Homo habilis en nuestro propio género y sería más gráfico denominarlo «Australopithecus habilis»: así el lector sabría mejor de qué clase de homínido estamos hablando. Sin embargo, no está tan claro que su mente fuera la de un australopiteco, la mente de un mono en definitiva.
En primer lugar se nota en Homo habilis una cierta expansión del cerebro con respecto de los australopitecos. Un cráneo procedente del lago Turkana es el que tiene un menor cerebro, de sólo 510 cc. Este cráneo en realidad se diferencia poco del de algunos Australopithecus africanus de Sterkfontein, aunque en algunos detalles muy pequeños, sutiles pero, sin embargo, importantes, se revela más cercano a nosotros. Otros cuatro cráneos de Homo habilis tienen cerebros algo mayores, de 582 cc, 594 cc, 638 cc y 674 cc (el primero procede del lago Turkana y los tres últimos de Olduvai). Existen no obstante grandes dudas acerca de cómo se han reconstruido los cerebros de estos fósiles, que están muy incompletos o deformados, y es probable que en los próximos años las estimaciones sufran un importante recorte en centímetros cúbicos.
Los cerebros como tales no se conservan, pero sí su forma y dimensiones, ya que queda el hueco en el interior del cráneo, el endocráneo, donde se alojaba el cerebro; mejor sería decir el encéfalo, porque aunque la mayor parte del endocráneo corresponde al cerebro, también están ahí el cerebelo y el bulbo raquídeo. Los paleontólogos rellenan el hueco endocraneal con escayola o con silicona o látex, y así se tiene una copia del encéfalo de un homínido de hace varios millones de años. De hecho es el único órgano del cuerpo que, aunque sea en forma de negativo, fosiliza.
Para complicar la taxonomía —es decir, la clasificación en especies— de los primeros fósiles del género Homo, Richard Leakey encontró un cráneo que no encaja fácilmente en esta historia. Richard Leakey es el descubridor de muchos fósiles famosos en la región del lago Turkana, y además marido de Meave Leakey, de quien también hemos hablado antes. Los padres de Richard, Louis y Mary, fueron los iniciadores de los hallazgos de fósiles humanos en el este de África, y obtuvieron éxitos resonantes en la garganta de Olduvai, en Tanzania. El cráneo que viene a complicarnos el relato lleva la sigla KNM-ER 1470, y su capacidad craneal es de nada menos que 752 cc. Podría tratarse de un gran macho de Homo habilis, si consideramos que esta especie tenía una gran diferencia entre los dos sexos, un gran dimorfismo sexual, como el de los australopitecos o aún mayor. Hay un problema para aceptar esta hipótesis, y es que la diferencia entre el KNM-ER 1470 y los demás fósiles de Homo habilis no reside sólo en el tamaño, sino también en la forma. El Homo habilis presenta una cara y unas muelas menores que las del Australopithecus africanus: en estos rasgos se acerca a nosotros. Sin embargo, lo que caracteriza al KNM-ER 1470, aparte de su gran cerebro, es su inmensa cara y su gran aparato masticador (una extraña combinación de rasgos). Por este motivo algunos autores consideran al KNM-ER 1470 y a algunos otros fósiles de grandes muelas (como la mandíbula de Malawi antes mencionada) una especie aparte (Homo rudolfensis).
El Homo habilis era un animal distinto de los anteriores homínidos también en términos ecológicos. Es la primera especie que no está completamente ligada a un medio forestal, ya sea una selva húmeda, caso del Ardipithecus ramidus, o un bosque más seco y menos denso, como los australopitecos. El Homo habilis parece haber habitado paisajes mucho más abiertos, como sabanas con árboles y matorrales, bien dispersos o bien formando manchas entre grandes claros de formaciones herbáceas. Este cambio ecológico es de la máxima importancia, porque abre la puerta a cambios aún mayores que vendrán más tarde, y que harán posible que los descendientes del Homo habilis acaben viviendo en toda clase de regiones, climas y ecosistemas. Sin excepción, los demás miembros de nuestro grupo de primates (gibones, orangutanes, gorilas y chimpancés), y todos nuestros antepasados hasta ese momento son o eran habitantes del bosque.
El cambio de hábitat que se produce con el Homo habilis coincide en el tiempo con un gran cambio climático, y puede ser su consecuencia. En los últimos cuatro millones de años el planeta viene experimentando una continua tendencia a hacerse cada vez más frío y seco. Alrededor de esa tendencia general se producen oscilaciones climáticas, bandazos térmicos que enfrían y calientan la Tierra periódicamente, al tiempo que la secan unas veces y la humedecen otras. Estas oscilaciones dependen de factores astronómicos, es decir, de los movimientos del eje de la Tierra y de la órbita que el planeta describe alrededor del Sol. Tales cambios astronómicos siguen determinados ciclos que afectan a la radiación solar que llega a la Tierra, y determinan los ciclos climáticos (junto con otros factores que no son orbitales).
El caso es que hasta hace 2,8 millones de años las oscilaciones climáticas se producían cada 23 000 años, aproximadamente, y eran de baja amplitud: no cambiaban mucho las cosas. Hace 2,8 millones de años empezaron a producirse oscilaciones cada 41 000 años, y esta vez eran de una amplitud mucho más considerable, de modo que en las épocas frías se acumulaban grandes masas de hielo alrededor de los polos. Además, posiblemente los dos casquetes polares, el ártico y el antártico, ya eran permanentes, y se mantenían, aunque menguados, en las épocas cálidas. Estos periódicos enfriamientos y desecaciones del planeta parecen haber tenido un enorme impacto ecológico en todas partes, incluyendo las regiones africanas donde vivían los homínidos, de manera que la selva lluviosa se redujo drásticamente y los ecosistemas abiertos se extendieron a su costa. La expansión de las sabanas, y el cambio vegetal que supuso, se vio acompañada por la evolución de varias líneas de mamíferos, que produjeron especies adaptadas al nuevo medio. Y entre ellas estaba el Homo habilis.