9
IL SÉ AUTOBIOGRAFICO
LA MEMORIA DIVENTA COSCIENTE
Le autobiografie sono costituite di ricordi personali, la somma totale delle esperienze della nostra vita, comprese quelle dei progetti che abbiamo fatto per il futuro, specifici o vaghi che siano. Il sé autobiografico - un’autobiografia resa cosciente - attinge dall’intero orizzonte della nostra storia memorizzata, remota e recente. In quella storia, sono comprese le esperienze sociali di cui siamo stati parte, o di cui avremmo voluto far parte; vi sono ugualmente compresi i ricordi che descrivono le nostre esperienze emozionali più raffinate, e precisamente quelle che hanno i requisiti per essere definite spirituali.
Mentre il sé nucleare pulsa incessantemente, sempre «Online», passando da un accenno appena abbozzato a una presenza manifesta, il sé autobiografico conduce una doppia vita. A un estremo, può essere esplicito, dando luogo alla mente cosciente nella sua espressione più alta e profondamente umana; all’altro estremo, può rimanere dormiente, con le sue innumerevoli componenti in attesa del proprio turno per attivarsi. Questa seconda vita del sé autobiografico si svolge in privato, lontano dalla coscienza accessibile, e forse è proprio lì - in termini di luogo e di tempo - che il sé matura grazie alla progressiva sedimentazione e alla rielaborazione della memoria. Nel momento in cui le esperienze vissute sono ricostruite e ri-esperite, sia nel corso della riflessione cosciente, sia in una elaborazione non cosciente, la loro sostanza viene rivalutata e inevitabilmente riorganizzata, e subisce modificazioni che, in termini di composizione fattuale e accompagnamento emozionale, possono essere minime o molto significative. Durante questo processo, entità ed eventi acquistano un nuovo peso emozionale. In sala di montaggio, alcuni fotogrammi della rievocazione sono lasciati cadere sul pavimento, altri vengono rigenerati e migliorati, e altri ancora sono combinati così abilmente dalla nostra volontà o dai capricci del caso da creare scene nuove, che non sono mai state girate. Ecco perché, con il passare degli anni, la nostra storia viene impercettibilmente riscritta. Ecco perché i fatti possono acquistare un nuovo peso e perché oggi la musica della memoria è diversa da quella dell’anno scorso.
In termini neurologici, questo lavoro di costruzione e ricostruzione si svolge in larga misura come elaborazione non cosciente e, per quello che ne sappiamo, forse ha luogo anche nei sogni; in qualche caso, tuttavia, può affiorare a livello della coscienza. Esso si avvale dell’architettura delle zone di convergenza-divergenza per trasformare la conoscenza criptata, presente nello spazio delle disposizioni, nelle rappresentazioni esplicite e decrittate contenute nello spazio delle immagini.
Considerata l’abbondanza delle registrazioni riguardanti il nostro passato vissuto e il nòstro futuro anticipato, è una fortuna che non occorra rievocarle tutte, e nemmeno la maggior parte, ogni volta che il nostro sé opera nella modalità autobiografica. Nemmeno Proust avrà dovuto attingere a tutto il suo remoto passato fitto di dettagli per costruire un istante del suo sé maturo in tutta la sua qualità proustiana. Invece - ed è un gran sollievo - facciamo affidamento su alcuni episodi chiave (in realtà su una raccolta di episodi) : a seconda delle esigenze del momento, ci limitiamo a rievocarne alcuni, in quanto rilevanti rispetto a quello nuovo. In certe situazioni, il numero degli episodi richiamati può essere elevatissimo, un’autentica inondazione di ricordi permeati delle emozioni e dei sentimenti originariamente associati agli episodi stessi. (Si può sempre contare su Bach per indurre qualcosa del genere). Ma anche quando il numero degli episodi è limitato, la complessità degli elementi da ricordare implicati nella strutturazione del sé è a dir poco enorme, ed è proprio qui che risiede il problema della costruzione del sé autobiografico.
LA COSTRUZIONE DEL SÉ AUTOBIOGRAFICO
Io sospetto che la strategia usata dal cervello per la costruzione del sé autobiografico sia la seguente. In primo luogo, occorre raggruppare insiemi significativi di ricordi biografici definitori, in modo che ciascuno di essi possa essere facilmente trattato come un singolo oggetto. Ognuno di tali oggetti può modificare il proto-sé e produrre una sua pulsazione del sé nucleare, dando luogo, a ruota, ai rispettivi sentimenti di conoscenza e alla prominenza conseguentemente accordata all’oggetto stesso. In secondo luogo, poiché gli oggetti presenti nella nostra biografia sono numerosissimi, il cervello necessita di dispositivi in grado di coordinare l’evocazione dei ricordi, di consegnarli al proto-sé per la necessaria interazione, e di conservare i risultati di quest’ultima in una configurazione coerente connessa agli oggetti causativi. Non si tratta di un problema banale; in effetti, i livelli complessi del sé autobiografico - quelli che, per esempio, riguardano importanti aspetti sociali - comprendono un numero così grande di oggetti biografici da richiedere numerose pulsazioni del sé nucleare. Di conseguenza, per costruire il sé autobiografico occorre un apparato neurale in grado sia di procurarsi molteplici pulsazioni del sé nucleare all’interno di una breve finestra temporale e per un gran numero di componenti, sia di tenere temporaneamente insieme i risultati di tali operazioni.
Da un punto di vista neurale, il processo di coordinamento è complicato soprattutto dal fatto che le immagini costituenti un’autobiografia sono in larga misura realizzate - sulla base della rievocazione di informazioni archiviate nelle cortecce disposizionali - nello spazio di lavoro delle immagini, ovvero nella corteccia cerebrale; ciò nondimeno, per essere rese coscienti, quelle stesse immagini devono interagire con l’apparato del proto-sé, il quale, come abbiamo visto, è in larga misura localizzato nel tronco encefalico. La costruzione di un sé auto-biografico richiede dunque meccanismi di coordinamento molto raffinati, dei quali la costruzione del sé nucleare può invece, in linea di massima, fare a meno.
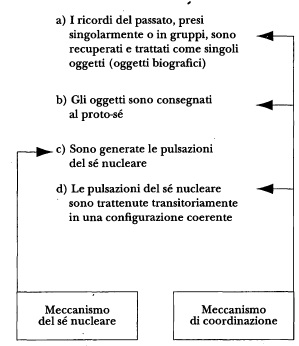
Figura 9.1 II sé autobiografico: meccanismi neurali.
Quale ipotesi di lavoro, allora, si può affermare che la costruzione del sé autobiografico dipende da due meccanismi congiunti. Il primo di essi è subordinato a quello del sé nucleare e garantisce che ciascun insieme di ricordi autobiografici sia trattato come un singolo oggetto e reso cosciente in una pulsazione del sé nucleare. Il secondo meccanismo mette a segno un’operazione di co-ordinamento che interessa tutto il cervello e che prevede i seguenti passaggi: 1) alcuni contenuti sono evocati dalla memoria e rappresentati come immagini; 2) le immagini possono interagire in modo ordinato con un altro sistema - precisamente il proto-sé - localizzato altrove nel cervello; e 3) i risultati dell’interazione sono tenuti insieme in modo coerente all’interno di una particolare finestra temporale.
Le strutture coinvolte nella costruzione del sé auto-biografico comprendono tutte quelle necessarie per il sé nucleare - localizzate nel tronco encefalico, nel talamo e nella corteccia cerebrale -, oltre a quelle coinvolte nei meccanismi di coordinamento discussi qui sotto.
IL PROBLEMA DEL COORDINAMENTO
Prima di dire una parola di più sul coordinamento, vorrei accertarmi di non essere frainteso. I dispositivi di coordinamento che sto ipotizzando non sono teatri cartesiani (al loro interno non è allestita alcuna rappresentazione). Non si tratta di centri della coscienza (non esiste nulla di simile). Non sono omuncoli interpreti (non hanno alcuna conoscenza e non interpretano niente). Sono esattamente quello che sto ipotizzando che siano, e nulla più. Sono organizzatori spontanei di un processo.
I risultati dell’intera operazione non si materializzano all’interno dei dispositivi di coordinamento, bensì altrove, e specificamente all’interno delle strutture del cervello che formano le immagini e generano la mente, localizzate nella corteccia cerebrale e nel tronco encefalico.
Il coordinamento non è guidato da un qualche misterioso agente esterno al cervello, ma da fattori naturali: per esempio, l’ordine con cui i contenuti, sotto forma di immagini, vengono immessi nel processo della mente, e il valore che viene loro riconosciuto. Come avviene la valutazione? Ogni immagine elaborata dal cervello è automaticamente stimata e associata a un valore, in un processo fondato sulle disposizioni: sia quelle originali del cervello (il suo sistema di valore biologico), sia quelle acquisite mediante l’apprendimento nell’arco di tutta la vita. La marcatura in base al valore - aggiunta durante la percezione originale e registrata insieme all’immagine -è ravvivata a ogni rievocazione. In breve, di fronte a certe sequenze di eventi e a una gran messe di conoscenza passata, filtrata e marcata in base al valore, i dispositivi di co-ordinamento del cervello contribuiscono organizzando i contenuti attuali. Inoltre, i dispositivi di coordinamento consegnano le immagini al sistema del proto-sé e infine trattengono i risultati dell’interazione (pulsazioni del sé nucleare) in una configurazione transitoria coerente.
I COORDINATORI
Nell’ipotesi di lavoro qui presentata, il primo stadio della realizzazione del sé autobiografico, a livello neurale, richiede strutture e meccanismi che sono già stati discussi per il sé nucleare. Vi è tuttavia qualcosa di distintivo nelle strutture e nei meccanismi necessari a realizzare il secondo stadio del processo, e in particolare il coordinamento già descritto che interessa il cervello nella sua totalità.
Quali sono i candidati a questo ruolo di coordinamento su ampia scala? Vengono in mente diverse strutture possibili, delle quali tuttavia solo alcune possono essere prese seriamente in considerazione. Un candidato importante è il talamo, in particolare l’insieme dei suoi nuclei associativi: una presenza costante in qualsiasi discussione sulle basi neurali della coscienza. La posizione dei nuclei talamici, intermedia fra la corteccia cerebrale e il tronco encefalico, è ideale per la gestione e il coordinamento dei segnali. Sebbene sia già abbastanza impegnato a costruire il tessuto di fondo di qualsiasi immagine, quando si tratta di coordinare i contenuti che definiscono il sé autobiografico, il talamo associativo ha un ruolo molto importante, anche se forse non il più importante in assoluto. Ritornerò sul tema del talamo e del coordinamento nel prossimo capitolo.
Quali sono le altre possibili candidature? Ve n’è una, che appare molto convincente, ed è quella di un insieme composito di regioni, presenti in entrambi gli emisferi, le cui connessioni mostrano un’architettura caratteristica. Ciascuna regione è un nodo macroscopico localizzato in corrispondenza di un importante incrocio tra vie di segnalazione convergente e divergente. Nel capitolo 6 le ho descritte come regioni di convergenza-divergenza (rcd) e ho specificato che ciascuna di esse è costituita da numerose zone di convergenza-divergenza. Le rcd sono localizzate strategicamente all’interno delle cortecce associative di ordine superiore, ma non all’interno delle cortecce sensoriali che creano le immagini. Esse si materializzano in siti quali la giunzione temporoparietale, le cortecce temporali laterali e mediali, le cortecce parietali laterali, le cortecce frontali laterali e mediali, e le cortecce posteromediali. Queste RCD conservano la registrazione di conoscenze acquisite in precedenza riguardanti i temi più vari; l’attivazione di una qualsiasi di queste regioni promuove la ricostruzione - per mezzo della divergenza e della retroattivazione nelle aree che creano
immagini - di vari aspetti della conoscenza passata, compresi quelli pertinenti alla biografia e quelli che descrivono conoscenze genetiche, non personali.
È plausibile che le principali rcd siano ulteriormente integrate da connessioni cortico-corticali a lungo raggio del tipo identificato da Jules Déjérine un secolo fa; tali connessioni introdurrebbero un ulteriore livello di co-ordinamento fra aree diverse.
Una delle principali rcd, la corteccia posteromediale (cpm), sembra essere, da un punto di vista funzionale,

Figura 9.2 II compito di coordinare le varie immagini generate dai processi di percezione e rievocazione è coadiuvato dalle regioni di convergenza-divergenza (rcd), localizzate all'interno delle cortecce associative non mappate. La localizzazione approssimativa delle principali rcd è indicata nei disegni (aree ombreggiate più scure) : cortecce temporali polare e mediale, corteccia prefrontale mediale, giunzioni temporoparietali e corteccia posteromediale (cpm). Con ogni probabilità vi sono altre regioni simili. La maggior parte delle rcd rappresentate nella figura fa anche parte della « rete di default» di Raichle discussa in seguito in questo capitolo. Per l'architettura di queste regioni, si vedano il capitolo 6 e le figure 6.1 e 6.2. Per i dettagli sulle connessioni di una regione di convergenza-divergenza, la cpm, si veda la figura 9.4.
gerarchicamente superiore rispetto alle altre, dalle quali si distingue per diversi tratti anatòmici e funzionali. Dieci anni fa ipotizzai che la regione cpm fosse legata al processo del sé, sebbene non nel ruolo che immagino adesso. Le informazioni ottenute negli ultimi anni hanno indicato l’effettivo coinvolgimento di questa regione nella coscienza, e in modo, specifico nei processi legati al sé; hanno inoltre fornito dati in precendenza non disponibili riguardanti la sua neuroanatomia e la sua fisiologia. (Queste informazioni sono discusse nelle parti finali del capitolo).
L’ultimo candidato è poco conosciuto, ma potrebbe rivelarsi vincente: è il claustro, una struttura misteriosa, strettamente legata alle rcd. Situato fra la corteccia dell’insula e i gangli basali di ciascun emisfero, il claustro ha connessioni corticali che potrebbero avere un potenziale ruolo di coordinamento. Francis Crick era convinto che il claustro fosse una sorta di regista delle operazioni sensoriali responsabili di legare fra loro le diverse componenti di un percetto multisensoriale. Le evidenze derivanti dalla neuroanatomia sperimentale rivelano effettivamente la presenza di connessioni con varie regioni sensoriali, il che rende perfettamente plausibile un suo ruolo di coordinamento. Fatto interessante, il claustro ha una robusta proiezione verso la cpm, l’importante regione di convergenza-divergenza già menzionata. La scoperta di questo forte legame ha avuto luogo solo dopo la morte di Crick, e quindi non è stata inclusa nell’articolo, pubblicato postumo, che egli scrisse con Christof Koch e in cui aveva esposto le sue posizioni.1 Quando si candida il claustro al ruolo di coordinatore, considerando il lavoro da eseguire, il problema sta nella piccola scala di questa struttura. D’altro canto, giacché non dovremmo aspettarci che nessuna delle strutture qui discusse svolga il lavoro di coordinamento in esclusiva, non vi è ragione perché il claustro non possa dare un importante contributo alla costruzione del sé autobiografico.
UN POSSIBILE RUOLO DELLA CORTECCIA POSTEROMEDIALE
Per determinare il ruolo specifico della cpm nella costruzione della coscienza occorrono ulteriori ricerche. Più avanti, in questo stesso capitolo, analizzerò i dati provenienti da diverse fonti: le ricerche sull’anestesia, sul sonno e su alcune condizioni neurologiche (che spaziano dal coma e dallo stato vegetativo alla malattia di Alzheimer), insieme agli studi sui processi legati al sé condotti con tecniche di neuroimmagine funzionale. Prima, però, esaminiamo i dati sulla cpm che appaiono più solidi e interpretabili, e cioè quelli derivanti dalla neuroanatomia sperimentale. Farò qualche ipotesi sui possibili meccanismi della cpm e sulle ragioni per le quali questa regione dovrebbe essere studiata.
Quando proposi che la cpm avesse un ruolo nel generare la soggettività, dietro a quell’idea vi erano due ragionamenti. Il primo riguardava il comportamento e il presunto stato mentale dei pazienti neurologici con un danno focale localizzato in questa regione, per esempio il danno associato agli stadi terminali della malattia di Alzheimer, o anche a casi estremamente rari di ictus e metastasi cerebrali. L’altro ragionamento si rifaceva alla ricerca teorica di una regione cerebrale fisiologicamente adatta a unire le informazioni riguardanti tanto l’organismo, quanto gli oggetti e gli eventi con i quali esso interagisce. La cpm era uno dei miei candidati, giacché pareva essere localizzata in corrispondenza di un’intersezione delle vie nervose coinvolte nel traffico delle informazioni provenienti dai visceri (enterocettive), dal sistema muscoloscheletrico (propriocettive e chinestetiche) e dal mondo esterno (esterocettive). Gli aspetti fattuali del quadro non sono in discussione, ma non vedo più la necessità del ruolo funzionale che avevo prospettato a suo tempo. Nondimeno, l’ipotesi stimolò indagini che produssero nuovi dati importanti.
Far progressi sulla base di quell’ipotesi non fu cosa facile; il problema principale stava nel fatto che le informazioni neuroanatomiche disponibili su questa regione erano molto limitate. Studi importanti avevano cominciato a descrivere la connettività di alcune parti della cpm,2 ma il suo schema di connessione generale non era stato studiato. In effetti, la regione non era ancora designata con un termine-ombrello, ma con quelli riferiti alle sue parti componenti: la corteccia del cingolo posteriore, la corteccia retrospleniale e il precuneo. La CPM, comunque la si voglia chiamare, non era ancora oggetto di attenzione come area cerebrale degna di particolare nota.
Per esplorare l’ipotesi che la CPM fosse implicata nella coscienza, fu necessario acquisire informazioni, prima non disponibili, sulla neuroanatomia delle sue connessioni. Per questa ragione, il nostro gruppo di ricerca intraprese uno studio di neuroanatomia sperimentale sui primati non umani; gli esperimenti furono eseguiti nel laboratorio di Josef Parvizi in collaborazione con Gary Van Hoesen. Lo studio - condotto su macachi - consisteva essenzialmente nel praticare numerose iniezioni di traccianti biologici in tutti i territori dei quali si doveva studiare la connettività neurale. Una volta iniettati in una data regione cerebrale, i traccianti biologici sono assorbiti dai singoli neuroni e trasportati lungo gli assoni fino alla destinazione naturale di questi ultimi, quale che sia la struttura a cui il neurone è connesso: questi sono i cosiddetti traccianti anterogradi. Un altro tipo di tracciante biologico, il tipo retrogrado, viene assorbito dai terminali assonici e trasportato in senso inverso: dal luogo in cui si trovano i terminali, qualunque esso sia, fino al corpo cellulare del neurone, nel suo sito di origine. Lo spostamento del tracciante consente di individuare, per ciascuna regione bersaglio, i siti di origine delle connessioni che la regione stessa riceve, come pure i siti ai quali la regione studiata invia i suoi messaggi.
La cpm è costituita da diverse sottoregioni. (Nella mappa citoarchitettonica di Brodmann, si tratta delle
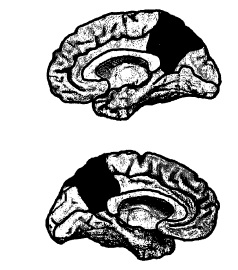
Figura 9.3 Localizzazione della corteccia posteromediale nel cervello umano. In alto, emisfero destro; in basso, emisfero sinistro.
aree 23a/b, 29, 30, 31 e 7m). L’interconnettività di queste sottoregioni è talmente intricata che, in una certa misura, è ragionevole trattarle come un’unità funzionale. I particolari rapporti di connessione all’interno dei sottosettori svelano la possibilità che alcune di esse abbiano ruoli funzionali distinti. Il termine ombrello che abbiamo coniato per l’ensemble sembra comunque giustificato, almeno per adesso.
Le connessioni della cpm, così come furono descritte nella prima pubblicazione scaturita da queste indagini difficili e assai dispendiose in termini di tempo,3 sono mostrate schematicamente nella figura 9.4 e possono essere spiegate come segue:
1. Gli input provenienti dalle cortecce associative temporale e parietale, come pure dalle cortecce entorina-
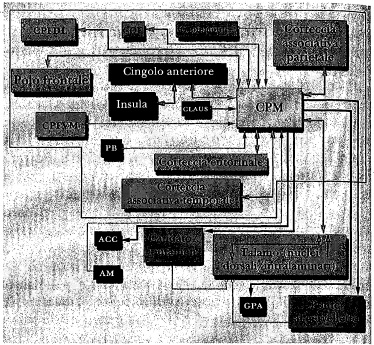
Figura 9.4 Le connessioni neurali da e verso la corteccia posteromediale (cpm) , determinate in uno studio condotto nelle scimmie. Abbreviazioni: c. = corteccia; cpfdl = corteccia prefrontale dorsolaterale; cof = campi oculari frontali; cpfvm = corteccia prefrontale ventromediale; pb = prosencefalo basale; claus = claustro; acc = nucleo accumbens; am = amigdala; gpa = grigio periacqueduttale.
le e frontale, convergono sulla cpm insieme a quelli provenienti dalla corteccia del cingolo anteriore (una delle principali regioni riceventi proiezioni dall’insula), dal claustro, dal prosencefalo basale, dall’amigdala, dalla regione premotrice e dai campi oculari frontali. I nuclei talamici intralaminari e dorsali proiettano anch’essi alla CPM.
2. Con poche eccezioni, i siti da cui originano gli input convergenti sulla cpm ricevono da essa, a loro volta, output divergenti: rappresentano un’eccezione, in tal senso, la corteccia prefrontale ventromediale, il claustro e i nuclei intralaminari del talamo. Vi sono poi alcuni siti che non proiettano alla cpm ma che ricevono le sue proiezioni: si tratta, precisamente, del caudato, del putamen, del nucleus accumbens e del grigio periacquedùttale.
3. Non vi sono connessioni dirette dalla cpm alle cortecce sensoriali di ordine inferiore o alle cortecce motrici primarie; né vi sono connessioni dirette da queste cortecce alla cpm.
4. Dai risultati descritti al punto 1 e 2, è evidente che la cpm è una regione di convergenza-divergenza di alto livello. Si tratta di un membro importante del gruppo delle rcd che io considero candidati promettenti al ruolo di coordinatore dei contenuti della mente cosciente; essa ha inoltre un’importante connessione con un altro potenziale coordinatore, il claustro, che proietta in misura significativa alla cpm ma riceve scarse afferenze reciproche.
Uno studio recente condotto sugli esseri umani ha corroborato l’idea che la cpm sia neuroanatomicamen-te distinta.4 Lo studio, diretto da Olaf Spoms, ha fatto uso di una moderna tecnica di risonanza magnetica, la dsi (Diffusion Spectrum Imaging), che produce immagini delle connessioni neurali e della loro approssimativa distribuzione spaziale. Gli autori hanno usato i dati così ottenuti per costruire mappe che mostrano l’organizzazione delle connessioni in tutta la corteccia cerebrale umana; vi hanno così identificato diversi hub - siti ad alta connettività -, molti dei quali corrispondono alle rcd che ho discusso fin qui. Gli autori dello studio hanno inoltre concluso che la regione cpm costituisce un hub con caratteristiche uniche, giacché intrattiene con altri hub rapporti più stretti di qualsiasi altro sito ad alta connettività.
LA CPM ALL’OPERA
Ci troviamo ora in una posizione migliore per immaginare come la cpm possa contribuire alla creazione della mente cosciente. Sebbene rappresenti una porzione considerevole della corteccia cerebrale, il potere della cpm non risiede tanto nei suoi possedimenti territoriali, quanto nelle sue relazioni. La cpm riceve segnali - e in larga misura ricambia la cortesia - dalla maggior parte delle regioni sensoriali associative di ordine superiore e dalle regioni premotrici. Le aree cerebrali ricche di zone di convergenza-divergenza, che hanno la chiave per accedere a insiemi compositi di informazione multimodale, sono pertanto in grado di segnalare alla cpm e in linea di massima possono ricevere a loro volta i suoi segnali. La cpm riceve anche segnali da nuclei sottocorticali coinvolti nello stato di veglia e a sua volta invia segnali a numerose regioni sottocorticali implicate nell’attenzione e nella ricompensa (a livello del tronco encefalico e del prosencefalo basale), come pure a regioni in grado di produrre routine motorie (per esempio i gangli basali e il grigio periacqueduttale).
Che cosa riguardano, probabilmente, tutti questi segnali in entrata? E che cosa ne fa la cpm? Non lo sappiamo per certo, ma l’enorme sproporzione fra la profusione e l’intensità delle proiezioni dirette alla cpm e le effettive dimensioni del suo territorio suggerisce una risposta: la cpm è perlopiù un’eredità antica, un territorio che si penserebbe impegnato nella gestione delle disposizioni, più che delle mappe esplicite: non è una moderna corteccia sensoriale di ordine inferiore, come quelle della visione o dell’udito, dove possono essere assemblate mappe dettagliate di oggetti ed eventi. Possiamo dire che la galleria della cpm non ha abbastanza spazio, sulle pareti, per esporre grandi dipinti oppure, se vogliamo, per allestire spettacoli di marionette. D’altra parte, va bene lo stesso, giacché anche le cortecce che inviano segnali alla cpm non sono come le cortecce sensoriali di ordine inferiore: anch’esse non possono esporre grandi dipinti né allestire spettacoli di marionette più di quanto possa farlo la cpm, e anch’esse sono in larga misura disposizionali; proprietarie di zone di convergenza-divergenza contenenti informazioni registrate.
Vista la sua organizzazione, la cpm - nel suo complesso e insieme ai suoi sottomoduli componenti - probabilmente si comporta essa stessa come una regione di con-vergenza-divergenza. Io immagino che l’informazione controllata dalla cpm e dai suoi partner possa essere riprodotta solo re-inviando segnali ad altre rcd del gruppo, le quali a loro volta possono inviare segnali alle cortecce sensoriali di ordine inferiore. Queste sono le cortecce in cui le immagini possono essere create e presentate, ovvero i siti in cui è possibile esporre grandi tele e allestire spettacoli di marionette. Rispetto ad altre regioni di convergenza-divergenza con le quali è interconnessa, la cpm ha uno speciale rango gerarchico: si trova più in alto sul totem, ed è in grado di dialogare in modo interattivo con le altre rcd.
In che modo, allora, la cpm contribuisce alla coscienza? Partecipando all’assemblaggio degli stati del sé auto-biografico. Ecco come l’immagino io: originariamente, attività sensoriali e motorie separate, legate all’esperienza personale, saranno state mappate nelle regioni cerebrali appropriate a livello corticale e sottocorticale, e i dati saranno stati registrati nelle zcd e nelle rcd. A sua volta, la cpm avrà costituito una rcd di ordine superiore interconnessa con le altre. Questa organizzazione avrà consentito all’attività all’interno della cpm di accedere a insiemi di dati più ampi e altamente distribuiti, con il vantaggio che il comando d’accesso sarebbe arrivato da un territorio relativamente piccolo e pertanto più gestibile dal punto di vista spaziale. La cpm poteva sostenere presentazioni di conoscenza momentanee e temporaneamente coese.
Se la connettività neuroanatomica della cpm è degna di nota, altrettanto lo è la sua localizzazione. La cpm è situata nei pressi della linea mediana, con la porzione sinistra che guarda la destra al di là del solco interemisferico. Questa posizione geografica nel volume cerebrale è conveniente per instaurare connessioni convergenti e divergenti con la maggior parte delle regioni del mantello corticale, ed è ideale per ricevere segnali dal talamo, e inviarne di rimando. Curiosamente, la localizzazione della CPM consente anche una protezione dagli impatti esterni; inoltre, essendo irrorata da tre vasi sanguigni importanti e indipendenti, la regione viene a essere relativamente immune nei confronti di danni vascolari o traumi che potrebbero distruggerla in modo radicale.
Come ho sottolineato in precedenza, le strutture che hanno a che fare con la coscienza condividono diversi tratti anatomici. In primo luogo, a livello corticale o sottocorticale esse tendono a essere di origine antica. Questo non dovrebbe sorprendere giacché, sebbene abbiano avuto luogo tardi nell’evoluzione biologica, gli esordi della coscienza non sono assolutamente uno sviluppo recente. In secondo luogo, tanto le strutture corticali, quanto quelle sottocorticali tendono a essere collocate sulla linea mediana o nei suoi pressi, e proprio come nel caso della cpm destra e sinistra, guardano le strutture gemelle dall’altra parte della linea mediana: questo accade nel caso dei nuclei talamici e ipotalamici, come pure in quello dei nuclei tegmentali del tronco encefalico. Qui, l’età evolutiva e la convenienza della localizzazione rispetto alla distribuzione diffusa dei segnali sono stretta-mente correlate.
Nei confronti della rete delle rcd corticali, la cpm opererebbe come un partner. Tuttavia, il ruolo delle altre rcd e l’importanza del sistema del proto-sé è tale che probabilmente, anche in seguito a un’ipotetica distruzione completa della regione cpm, la coscienza sarebbe compromessa ma non abolita purché tutte le altre rcd e il sistema del proto-sé rimanessero intatti. La coscienza sarebbe ripristinata, sebbene non al massimo grado. Nel caso degli ultimi stadi della malattia di Alzheimer, che descriverò nella prossima sezione, la situazione è diversa, nel sènso che l’insulto subito dalla cpm è virtualmente l’ultima goccia in un processo di graduale devastazione che ha già invalidato altre rcd e il sistema del proto-sé.
ALTRE CONSIDERAZIONI SULLA CORTECCIA POSTEROMEDIALE
Ricerche sull’anestesia
Per alcuni aspetti, l’anestesia generale è un mezzo ideale per indagare la neurobiologia della coscienza. Si tratta di uno degli sviluppi più spettacolari della medicina e ha salvato la vita di milioni di persone altrimenti inoperabili. Spesso si pensa all’anestesia generale come a un analgesico, perché i suoi effetti impediscono di avvertire il dolore causato dalle ferite chirurgiche. La verità, però, è che l’anestesia preclude il dolore nel modo più radicale possibile; essa infatti sospende del tutto la coscienza: non soltanto il dolore, ma tutti gli aspetti della mente cosciente.
Un’anestesia superficiale riduce leggermente la coscienza lasciando spazio, in una certa misura, all’apprendimento inconscio e all’occasionale «affiorare» dell’elaborazione cosciente. L’anestesia profonda, d’altra parte, penetra completamente nel processo della coscienza e, a tal proposito, costituisce una variazione farmacologicamente controllata dello stato vegetativo o addirittura del coma. Questo è esattamente ciò che serve a un chirurgo per lavorare indisturbato su un cuore o su un’anca. Il paziente deve essere lontano, lontanissimo dal tavolo operatorio, incapace di muoversi e così profondamente addormentato che i suoi muscoli hanno il tono della gelatina. L’anestesia necessaria per operare è quella dello stadio III: il paziente non ode nulla, non sente nulla e non pensa a nulla. Se il chirurgo gli rivolge la parola, non risponde.
Nel corso della storia, l’anestesia ha messo a disposizione dei chirurghi numerosi agènti farmacologici con cui lavorare, e la ricerca di molecole che funzionino nel modo più efficace presentando una scarsa tossicità e comportando rischi minimi è tuttora in corso. In linea di massima, gli anestetici svolgono il loro compito aumentando l’inibizione a livello dei circuiti neurali. Questo si può ottenere rafforzando l’azione del gaba (acido gamma-aminobutirrico), che nel cervello è il principale trasmettitore ad azione inibitoria. Gli anestetici agiscono iperpo-larizzando i neuroni e bloccando l’acetilcolina, una molecola importante nella normale comunicazione inter-neuronale. Un tempo era convinzione comune che gli agenti anestetici esercitassero la loro azione deprimendo la funzione cerebrale in modo indiscriminato, riducendo l’attività dei neuroni praticamente ovunque. Studi recenti hanno invece dimostrato che alcuni anestetici agiscono in modo molto selettivo, esercitando la loro azione a livello di siti cerebrali specifici. Un esempio, a tal proposito, è quello del propofol. Come mostrano alcuni studi di neuroimmagine funzionale, esso produce il suo splendido effetto agendo principalmente su tre siti: la corteccia posteromediale, il talamo e il tegmento del tronco encefalico. Sebbene l’importanza relativa di ciascuno di questi siti nella produzione dell’incoscienza sia sconosciuta, la diminuzione del livello di coscienza è correlata alla riduzione del flusso ematico regionale nella corteccia posteromediale. Le evidenze, tuttavia, vanno ben oltre il propofol: altri agenti anestetici sembrano avere effetti paragonabili, come ha dimostrato una review di ampio respiro. Nell’anestesia indotta dal propofol sono selettivamente depressi tre territori cerebrali paramediani coinvolti nella creazione della coscienza.5
Ricerca sul sonno
Il sonno rappresenta un contesto naturale per lo studio della coscienza, e le ricerche che lo riguardano sono state infatti fra le prime a contribuire alla comprensione del problema. È ormai assodato che i ritmi elettroencefalografici - i tipici pattern di attività elettrica generati dal cervello - sono associati a stadi del sonno specifici. È notoriamente difficile ricondurre l’origine dei pattern elettroencefalografici a particolari regioni cerebrali, ed è proprio qui che la localizzazione spaziale consentita dalle tecniche di neuroimmagine funzionale si è rivelata utilissima per completare il quadro. Nell’arco degli ultimi dieci anni, esse hanno infatti consentito di osservare più da vicino, durante i vari stadi del sonno, regioni cerebrali specifiche.
La coscienza è profondamente depressa, per esempio, durante il sonno a onde lente, noto anche come sonno non rem, ossia caratterizzato da movimenti oculari non rapidi. Si tratta del sonno profondo, il classico sonno del giusto, quello dal quale può ridestarci soltanto una sveglia sgarbata e ingiusta. È il « sonno senza sogni », sebbene la completa assenza di questi ultimi sembri valere solo per la prima parte della notte. Gli studi di neuroimmagine funzionale dimostrano che in un certo numero di regioni cerebrali, durante il sonno a onde lente, l’attività è ridotta: ciò avviene soprattutto in alcune parti del tegmento del tronco encefalico (il ponte e il mesencefalo), nel diencefalo (il talamo e l’ipotalamo/prosen-cefalo basale), nelle parti mediali e laterali della corteccia prefrontale, nella corteccia del cingolo anteriore, nella corteccia parietale laterale, e nella cpm. Nel sonno a onde lente, la riduzione funzionale è meno selettiva rispetto a quanto osservato nell’anestesia generale (del resto non vi è ragione perché le modalità debbano essere le stesse) ; come nell’anestesia, però, non vi sono indicazioni di una depressione funzionale indiscriminata. Il fenomeno interessa in modo prominente le tre strutture legate alla creazione della coscienza (tronco encefalico, talamo e cpm), le quali risultano tutte e tre depresse.
La coscienza è depressa anche durante il sonno rem (con movimenti oculari rapidi), nel corso del quale sono assai frequenti i sogni. Il sonno rem, tuttavia, consente ai contenuti dei sogni di penetrare nella coscienza: o attraverso l’apprendimento e la successiva rievocazione, o attraverso la cosiddetta coscienza paradossa. Le regioni del cervello la cui attività è più marcatamente ridotta durante il sonno rem sono la corteccia prefrontale dorsolaterale e la corteccia parietale laterale; prevedibilmente, a livello della cpm, la riduzione di attività è molto meno marcata.6
In breve, il livello di attività della cpm è massimo durante lo stato di veglia e minimo durante il sonno a onde lente; durante il sonno rem la cpm opera a livelli intermedi. Questo è logico: nel sonno a onde lente la coscienza è in larghissima misura sospesa, mentre nel sonno rem gli eventi accadono a un « sé ». Certo, il sé del sogno non è il sé normale, tuttavia lo stato cerebrale associato sembra reclutare la cpm.
Il coinvolgimento della CPM nella rete di default
In una serie di studi di immagine funzionale che hanno fatto uso sia della tomografia a emissione di positroni, sia della risonanza magnetica funzionale, Marcus Raichle ha richiamato l’attenzione sul fatto che quando i soggetti si trovano in uno stato di riposo, non impegnati nell’esecuzione di compiti che richiedano un’attenzione concentrata, sembra essere costantemente attivato un particolare sottoinsieme di regioni cerebrali; al contrario, quando l’attenzione si appunta su un compito specifico, l’attività di queste regioni diminuisce leggermente; mai, però, al livello osservato per esempio nell’anestesia.7 L’insieme di queste regioni comprende la corteccia prefrontale mediale, la giunzione temporoparietale, alcune strutture della corteccia temporale anteriore e mediale, e la cpm: tutte regioni che oggi sappiamo essere estesamente interconnesse. In effetti, gran parte dell’attenzione concentrata sulla cpm è stata solle-

Figura 9.5 Nell’imaging funzionale, la cpm, insieme ad altre regioni di convergenza-divergenza, mostra una significativa attivazione durante l’esecuzione di diversi compiti autoreferenziali. Tali compiti comprendono il richiamo di ricordi autobiografici, l’anticipazione di eventi futuri e l’elaborazione di giudizi morali.
citata proprio dalla sua appartenenza a questo insieme di regioni.
Raichle ha ipotizzato che l’attività di questa rete rappresenti una modalità operativa «di default» che viene interrotta da compiti che richiedono un’attenzione diretta verso l’esterno. Nell’esecuzione di compiti che necessitano invece di un’attenzione rivolta verso l’interno e orientata sul sé (per esempio il richiamo di informazioni autobiografiche e certi stati emozionali), noi e altri abbiamo dimostrato che la riduzione dell’attività della cpm è meno pronunciata o può non apparire affatto. In tali condizioni, anzi, essa può alimentare.8 Esempi sono la rievocazione di ricordi autobiografici, la rievocazione di progetti per un futuro possibile, alcuni compiti legati alla « teoria della mente » e molti compiti che comportano la formulazione di giudizi su persone o situazioni all’interno di un contesto morale.9 In tutti questi compiti, tende a essere presente un altro importante sito di attività, anche se non altrettanto esteso: un altro territorio mediale, localizzato anteriormente nella corteccia prefrontale. Sappiamo che da un punto di vista neuroanatomico anche questa è una regione di convergenza-divergenza.
Raichlé ha enfatizzato l’aspetto intrinseco della modalità operativa di default e l’ha messo in relazione, molto ragionevolmente, con l’elevatissimo consumo di energia associato all’attività cerebrale intrinseca, in contrapposizione ad attività guidate da stimolazioni esterne: con ogni probabilità, la cpm è, in tutta la corteccia cerebrale, la regione che presenta il metabolismo più elevato.10 Anche ciò è compatibile con il ruolo che sto proponendo per questa regione nella creazione della coscienza, ovvero quello di un importante integratore/coordinatore che rimarrebbe attivo in ogni istante, cercando di trattenere in una configurazione coerente insiemi altamente disparati di attività di fondo. In che modo l’andamento fluttuante della modalità operativa di default è compatibile con l’idea che una regione come la CPM possa contribuire alla coscienza? Forse esso riflette la coreografia fra primo piano e sfondo, messa in atto dal sé all’interno della mente cosciente. Quando abbiamo bisogno di fare attenzione a stimoli esterni, la nostra mente cosciente porta l’oggetto in esame in primo piano e lascia che il sé si ritiri sullo sfondo. Quando non siamo sollecitati dal mondo esterno, il nostro sé si sposta invece verso il centro del palcoscenico e può anche venire in primo piano quando l’oggetto in esame è la persona stessa, da sola o in un contesto sociale.
Ricerche su alcune condizioni di interesse neurologico
L’elenco delle condizioni di interesse neurologico nelle quali la coscienza è compromessa è fortunatamente breve: il coma e gli stati vegetativi, alcuni tipi di stati epilettici, nonché i cosiddetti stati di mutismo acinetico che possono essere causati da alcuni tipi di ictus, da tumori e dagli stadi terminali della malattia di Alzheimer. Nel coma e negli stati vegetativi, la compromissione è radicale, come se un territorio cerebrale venisse intenzionalmente e brutalmente colpito con una mazza.
Malattia di Alzheimer. L’Alzheimer, una malattia esclu-sivamente umana, è anche uno dei più gravi problemi sanitari dei tempi moderni. Mentre facciamo i nostri tentativi di comprenderla, però, affiora anche un corollario in qualche modo positivo, giacché essa è diventata una fonte di preziose informazioni sulla mente, sul comportamento e sul cervello. Il contributo dell’Alzheimer alla comprensione della coscienza sta diventando chiaro soltanto ora.
A partire dagli anni Settanta, ho avuto l’opportunità di seguire molti pazienti colpiti da questa malattia e il privilegio di effettuare studi post mortem sul loro cervello effettuando esami sia a livello macroscopico, sia sul materiale microscopico. In quegli anni parte del nostro programma di ricerca era dedicato alla malattia di Alzheimer e il mio collega e stretto collaboratore Gary W. Van Hoesen era uno dei massimi esperti della neuroanatomia del cervello colpito da questa patologia. All’epoca, il nostro principale obiettivo era quello di comprendere in che modo l’alterazione dei circuiti cerebrali osservata nei soggetti con Alzheimer potesse causare il disturbo della memoria che caratterizza la loro condizione.
La maggior parte dei pazienti con la tipica malattia di Alzheimer non ha disturbi della coscienza: né durante i primi stadi della malattia, né a uno stadio intermedio. I primi anni di malattia sono contraddistinti da un progressivo venir meno della capacità di apprendere informazioni fattuali nuove e di rievocare quelle apprese in precedenza. Sono anche comuni difficoltà di giudizio e di orientamento spaziale. In una prima fase la malattia può colpire in modo così lieve che per qualche tempo il paziente può conservare le sue capacità sociali e una parvenza di vita normale.
Al principio degli anni Ottanta, il nostro gruppo di ricerca, che ora comprendeva anche Brad Hyman, individuò nelle estese alterazioni neuropatologiche a livello della corteccia entorinale e dei campi adiacenti della corteccia temporale anteriore una causa plausibile del deficit di memoria fattuale riscontrabile nella malattia di Alzheimer.11 L’ippocampo - la struttura cerebrale necessaria per fissare nuovi ricordi fattuali in altre aree del cervello - era a tutti gli effetti disconnesso dalle cortecce entorinale e temporale anteriore. Di conseguenza, non potevano essere appresi nuovi fatti. Inoltre, con il progredire della malattia, la corteccia del lobo temporale anteriore era essa stessa danneggiata a tal punto da impedire l’accesso a informazioni fattuali particolari precedentemente apprese. Il fondamento stesso della memoria autobiografica era eroso e infine annientato esattamente come nei pazienti con una estesa distruzione del lobo temporale causata dall’encefalite erpetica, un’infezione virale che compromette anche, in modo selettivo, le regioni temporali anteriori. La specificità cellulare della malattia di Alzheimer era singolare. I neuroni degli strati II e IV della corteccia entorinale erano trasformati - in gran parte, se non nella loro totalità - in altrettante pietre tombali: la migliore espressione per descrivere ciò che resta dopo che la malattia li ha ridotti a grovigli neurofibrillari. Questo insulto selettivo realizzava un taglio netto, quasi come fosse praticato con un rasoio, nelle vie dirette all’ippocampo, che si servono dello strato II come stazione di relè. E per rendere completa la separazione, l’insulto produceva anche un taglio ugualmente netto nelle vie in uscita dall’ippocampo, che usano invece lo strato IV. C’è poco da meravigliarsi, quindi, se la memoria fattuale dei pazienti con Alzheimer è devastata.
Con il progredire della malattia, però, insieme ad altri disturbi mentali selettivi comincia a soffrire anche l’integrità della coscienza. Inizialmente il problema è prevedibilmente confinato alla coscienza autobiografica. Poiché i ricordi riguardanti eventi personali del passato non possono essere correttamente rievocati, il legame fra eventi attuali e passato vissuto diventa inefficiente. La coscienza riflessiva nell’elaborazione deliberativa offline è compromessa. Con ogni probabilità, questo disturbo - in parte, sebbene forse non completamente - è ancora da attribuirsi alla disfunzione del lobo temporale mediale.
Ancora più avanti, nella sua marcia inesorabile, la devastazione si estende ben oltre i processi autobiografici. Negli ultimi stadi dell'Alzheimer - osservabili nei pazienti che ricevono cure mediche e assistenza di buona qualità e che quindi sopravvivono più a lungo -, si instaura a poco a poco uno stato virtualmente vegetativo. La connessione di questi pazienti con il mondo è ridotta a tal punto che essi fanno pensare agli individui con mutismo acinetico: è sempre più raro che inizino spontaneamente a interagire con l’ambiente umano e fisico che li circonda; rispondono sempre meno agli stimoli; le loro emozioni sono messe a tacere. Il comportamento è dominato da un’espressione assente, indifferente, vuota, non concentrata, silenziosa.
Come spiegare la svolta che ha luogo nell’ultima fase dell'Alzheimer? Una risposta definitiva non è possibile perché, nel corso degli anni, la patologia si esprime in diversi siti cerebrali, e le alterazioni non sono limitate ai grovigli neurofibrillari. In una certa misura, però, il danno rimane selettivo. Le parti del cervello implicate nella creazione di immagini, e precisamente le cortecce sensoriali - uditiva e visiva - di ordine inferiore, non sono gravate dalla malattia, né lo sono le regioni motrici nella corteccia cerebrale, nei gangli basali e nel cervelletto. D’altro canto, alcune regioni legate alla regolazione dei processi vitali, dalle quali dipende il proto-sé, sono progressivamente danneggiate; esse comprendono non solo la corteccia dell’insula, ma anche il nucleo parabrachiale (un altro dato, questo, accertato dal nostro gruppo di ricerca).12 Infine, mostrano gravi danni altri settori del cervello ricchi di regioni di convergenza-divergenza, fra cui spicca in modo particolare la cpm.
La ragione per la quale sto prestando una particolare attenzione a questi fatti è che nelle fasi precoci della malattia la cpm mostra soprattutto placche neuritiche, ma negli stadi terminali il quadro è dominato dalla deposizione di grovigli neurofibrillari: le pietre tombali di neuroni in precedenza sani, alle quali alludevo prima. La loro estesa presenza nella cpm indica che il funzionamento della regione è gravemente compromesso.13
Eravamo ben consapevoli delle importanti alterazioni patologiche a livello della cpm, alla quale all’epoca ci riferivamo semplicemente come alla « corteccia del cingolo posteriore e aree limitrofe ». La ripetuta osservazione clinica, negli stadi terminali dell'Alzheimer, di una compromissione della coscienza nei casi di danno focale a questa regione, unitamente alla sua peculiare localizzazione anatomica, mi spinse tuttavia a domandarmi se una cpm seriamente danneggiata non potesse essere la goccia che fa traboccare il vaso.14
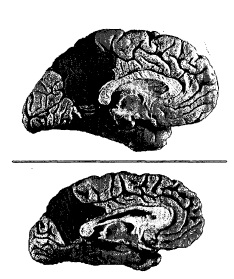
Figura 9.6 II disegno in alto mostra la superficie mediale dell’emisfero cerebrale sinistro in un individuo anziano normale. La cpm è ombreggiata. Il disegno in basso mostra la stessa superficie in un individuo approssimativamente della stessa età con malattia di Alzheimer a uno stadio avanzato. La cpm, anche qui ombreggiata, è gravemente atrofica.
Perché questa regione è un bersaglio della malattia di Alzheimer? La ragione potrebbe benissimo essere la stessa invocata da me e dai miei colleghi, molti anni or sono, per spiegare, sempre nell’Alzheimer, il coinvolgimento patologico diffuso delle regioni temporomediali.15 In condizioni di salute normali la corteccia entorinale e l’ippocampo non cessano mai di funzionare: lavorano giorno e notte per contribuire all’elaborazione di ricordi fattuali iniziando e consolidando la registrazione dei ricordi. Di conseguenza, la tossicità cellulare locale, associata a importanti fenomeni di usura, avrebbe un contraccolpo sui preziosi neuroni della regione. Lo stesso ragionamento potrebbe benissimo applicarsi alla cpm, visto il suo funzionamento pressoché continuo in una molteplicità di processi legati al sé.16
Riassumendo, i pazienti con Alzheimer terminale e un’evidente compromissione deella coscienza presentano un danno nauronale sproporzionato, e quindi una disfunzione, a livello di due territori cerebrali la cui integrità è necessaria per uno stato di coscienza normale: la cpm e il tegmento del tronco encefalico. Occorre essere prudenti circa l’interpretazione di questi fatti, giacché nell’Alzheimer vi sono anche altri siti di disfunzione. Allo stesso tempo, però, sarebbe sciocco non considerare queste evidenze.
Che cosa possiamo dire dei pazienti a quest’ultimo stadio della malattia, quando il loro cervello riceve un ulteriore colpo? In passato era stata mia opinione - e lo è tuttora — che, sebbene il nuovo insulto sia uno spettacolo doloroso per chi sta vicino alle vittime, probabilmente per il diretto interessato rappresenta una sorta di benedizione, sia pure sotto mentite spoglie. A questo stadio terminale, e con questo livello di compromissione della coscienza, i pazienti non possono assolutamente essere consapevoli dei danni arrecati dalla malattia. Di loro, ormai, non resta che l’involucro degli esseri umani che erano un tempo: di certo meritano di ricevere tutto il nostro amore e di essere accompagnati con le nostre cure alla loro triste fine. In una qualche misura, però, adesso sono misericordiosamente liberi dalle leggi del dolore e della sofferenza che ancora si applicano a chi li osserva.
Il coma, lo stato vegetativo e le differenze rispetto alla sindrome «locked-in»
In larga misura, i pazienti in coma non rispondono ai tentativi di comunicazione provenienti dal mondo esterno, sprofondati come sono in un sonno in cui spesso anche il ritmo della respirazione appare anormale. Non compiono gesti né emettono suoni significativi, meno che mai fanno uso di parole. Nessuna delle componenti fondamentali della coscienza elencate nel capitolo 8 è rilevabile. Lo stato di veglia è sicuramente assente; e, sulla base del comportamento che possiamo osservare, è ragionevole presumere che siano assenti pure la mente e il sé.
I pazienti in coma hanno spesso subito un danno al tronco encefalico, danno che a volte sconfina nell’ipotalamo; molto comunemente, tutto questo è causato da un ictus. Sappiamo che il danno deve essere localizzato nella parte posteriore del tronco encefalico, il tegmento, e più specificamente nella sua porzione superiore, che ospita i nuclei implicati nella regolazione dei processi vitali ma non quelli indispensabili per il mantenimento della funzione cardiorespiratoria. In altre parole, se il danno coinvolge anche la porzione inferiore del tegmento, il risultato non è il coma, ma la morte.
Quando il danno interessa la parte anteriore del tronco encefalico, ancora una volta il risultato non è il coma, ma la sindrome lockedrin, una condizione orribile in cui il paziente è assolutamente cosciente ma quasi del tutto paralizzato: può comunicare solo con ammiccamenti, a volte di un occhio soltanto, oppure con il movimento verso l’alto di un occhio. Nondimeno, questi pazienti vedono perfettamente qualsiasi cosa sia messa loro davanti, e quindi possono leggere. Possono anche udire benissimo e apprezzare il mondo nei minimi dettagli. La loro prigionia è pressoché completa; solo un certo ottundimento delle reazioni emozionali di fondo trasforma in qualche modo una situazione terrificante in una condizione a malapena tollerabile, per quanto dolorosa.
Conosciamo le esperienze uniche vissute da questi pazienti grazie ai resoconti che alcuni di loro, soggetti intelligenti e dotati di spirito d’osservazione, hanno avuto il coraggio di dettare, aiutati da esperti. In realtà, i loro resoconti non sono dettati, ma comunicati per mezzo dell'ammiccamento: una lettera, un battito di ciglia. Pensavo che la malattia di Lou Gehrig (la sclerosi laterale amiotrofica) fosse la patologia neurologica più crudele; si tratta di una condizióne cerebrale degenerativa, in cui i pazienti, anch’essi coscienti, perdono a poco a poco la capacità di muoversi, di parlare, e alla fine anche di deglutire. Dopo aver visto il mio primo paziente con la sindrome lockedrin, però, mi resi conto che questa condizione era addirittura peggiore. I due libri migliori scritti da pazienti locked-in sono documenti brevi e semplici, ma umanamente molto ricchi. Da uno di essi, scritto da Jean-Dominique Bauby, è stato tratto un film di sorprendente accuratezza, Lo scafandro e la farfalla, per la regia del pittore Julián Schnabel. Per i non specialisti si tratta di un documentario esauriente sulla sindrome.17
Il coma spesso sfuma in una condizione un poco più leggera, denominata stato vegetativo. Anche in questo caso il paziente è incosciente, ma come notato in precedenza lo stato vegetativo differisce dal coma per due motivi. In primo luogo, i pazienti presentano un’alternanza sonno-veglia, e quando si trovano nell’uno o nell’altro stato è presente anche un riscontro a livello elettroencefalografico; nella parte del ciclo corrispondente alla veglia, gli occhi possono essere aperti. In secondo luogo, i pazienti eseguono alcuni movimenti spontanei e possono rispondere con dei movimenti; non rispondono con la parola, però, e i movimenti eseguiti non sono mai specifici. Lo stato vegetativo può sfociare nel recupero della coscienza oppure rimanere stabile, nel qual caso è denominato « stato vegetativo persistente ». Oltre al danno interessante il tegmento del tronco encefalico e l’ipotalamo - che è la tipica patologia del coma -, lo stato vegetativo può scaturire da un danno al talamo e anche da un danno diffuso alla corteccia cerebrale o alla sostanza bianca sottostante.
Perché il coma e lo stato vegetativo hanno a che fare con il ruolo della cpm, visto che le lesioni causative sono localizzate altrove? Questa domanda è stata affrontata nel corso di studi di neuroimmagine funzionale che si proponevano di indagare in che misura le alterazioni funzionali che si verificano nel cervello dei pazienti in queste condizioni siano generalizzati o circoscritti. I soliti sospetti effettivamente non mancano di presentarsi, come rivela un’importante riduzione della funzione di strutture quali il tronco encefalico, il talamo e la cpm; tuttavia, la riduzione del tasso metabolico locale per il glucosio osservabile a livello della cpm è particolarmente pronunciata.18
Vi è peraltro un ulteriore risultato importante da riferire. Di solito, i pazienti in stato di coma o muoiono, o migliorano in modo molto modesto, passando a uno stato vegetativo persistente. Alcuni, però, sono più fortunati ed emergono gradualmente dal loro stato di coscienza profondamente compromessa. Nel momento in cui ciò accade, i cambiamenti più significativi a livello di metabolismo cerebrale hanno luogo nella cpm.19 Ciò indica una buona correlazione fra il livello di attività in quest’area e il livello di coscienza. Poiché la cpm ha un metabolismo tanto elevato, vi è la tentazione di liquidare questa osservazione come il risultato di un complessivo miglioramento dell’attività cerebrale. La cpm migliorerebbe per prima semplicemente a causa del suo elevato metabolismo. Ma questo non spiegherebbe perché, allo stesso tempo, venga recuperata la coscienza.
NOTA CONCLUSIVA SULLE PATOLOGIE DELLA COSCIENZA
Nello sforzo teso a delinearne la neuroanatomia, le patologie della coscienza hanno fornito importanti indicatori e hanno suggerito, in parte, i meccanismi proposti per la costruzione del sé nucleare e del sé autobiografico. Forse è utile concludere il capitolo stabilendo un legame trasparente fra la patologia umana da un lato e le ipotesi presentate in precedenza dall’altro.
Escludendo le alterazioni della coscienza che affiorano naturalmente durante il sonno, o che sono indotte dagli anestetici somministrati sotto il controllo medico, la maggior parte dei disturbi della coscienza deriva da una qualche profonda disfunzione cerebrale. In alcuni casi, il meccanismo è chimico: questo avviene in seguito al sovradosaggio di vari farmaci, compresa l’insulina somministrata per il trattamento del diabete, come pure nel caso di eccessivi livelli ematici di glucosio nel paziente diabetico non curato. L’effetto di queste sostanze chimiche è al tempo stesso selettivo e generalizzato. Con un intervento tempestivo e adeguato, tuttavia, la condizione è reversibile. D’altro canto, il danno strutturale causato da traumi cranici, da ictus o da certe malattie degenerative produce spesso disturbi della coscienza dai quali un completo recupero è un’evenienza improbabile. In alcune situazioni, poi, il danno cerebrale può anche dar luogo a crisi epilettiche: durante o dopo questi fenomeni, la presenza di stati alterati di coscienza è un sintomo importante.
I casi di coma e di stato vegetativo dovuti a danni del tronco encefalico presentano una compromissione tanto del sé nucleare, quanto del sé autobiografico. Essenzialmente, le principali strutture del proto-sé vengono o distrutte o gravemente danneggiate, così che non possono più essere generati né i sentimenti primordiali, né i « sentimenti di ciò che accade ». L’integrità del talamo e della corteccia cerebrale non è sufficiente a compensare il totale venir meno del sistema del sé nucleare. Tali condizioni testimoniano la precedenza gerarchica del sistema del sé nucleare e la completa dipendenza da quest’ultimo del sistema del sé autobiografico. È importante notare questo fatto, giacché non vale invece l’inverso: il sé autobiografico può essere compromesso in presenza di un sé nucleare altrimenti intatto.
I casi di coma o di stato vegetativo persistente in cui, invece di colpire il tronco encefalico, il danno compromette la corteccia, il talamo o le connessioni di queste strutture con il tronco encefalico possono causare una disfunzione del sé nucleare senza distruggerlo, il che spiega la progressione di alcuni di questi casi verso uno stato di coscienza «minima» e il recupero di alcune attività non coscienti legate alla mente. I casi di mutismo acinetico e di automatismo che si verificano dopo una crisi epilettica causano una compromissione reversibile del sistema del sé nucleare e una conseguente alterazione del sistema del sé autobiografico. In tali circostanze, sono comunque presenti alcuni comportamenti appropriati i quali, sebbene automatici, indicano che i processi mentali non sono del tutto aboliti.
Quando i disturbi del sé autobiografico appaiono in modo indipendente, in presenza di un sistema del sé nucleare altrimenti intatto, la causa va ricercata in qualche aspetto di una disfunzione mnemonica, un’amnesia acquisita. La causa più importante di amnesia è l’Alzhei-mer, che abbiamo appena visto; fra le altre cause vi sono l’encefalite virale e l’anossia acuta (interruzione dell’ossigenazione cerebrale) che può verificarsi durante un arresto cardiaco. Nei casi di amnesia, vi è una considerevole disgregazione dei ricordi particolari dell’individuo corrispondenti al suo passato e ai suoi progetti per il futuro. Ovviamente, i pazienti con un danno che interessi entrambe le regioni ippocampali-entorinali, la cui capacità di formare nuovi ricordi è compromessa, soffrono di una progressiva riduzione della portata del sé autobiografico, perché i nuovi eventi che si verificano nella loro vita non vengono correttamente registrati e integrati nella loro biografia. Più grave è la situazione dei pazienti il cui danno cerebrale investa non solo le regioni ippocampali-entorinali, ma anche quelle circostanti le cortecce entorinali, nel settore anteriore del lobo temporale. Questi pazienti sembrano perfettamente coscienti - il funzionamento del loro sé nucleare è intatto -, al punto che sono perfino consapevoli di non riuscire a ricordare. Tuttavia, la loro capacità di evocare la propria biografia, insieme a tutta l’informazione sociale a essa legata, è più o meno diminuita. Il materiale con cui è possibile assemblare un sé autobiografico è impoverito, o perché non può essere estratto dai ricordi del passato, o perché qualsiasi cosa venga estratta non può essere debitamente coordinata e consegnata al sistema del proto-sé (o forse per entrambi i motivi). Il caso estremo è quello del paziente B., il cui ricordo autobiografico - assolutamente schematico - è in larga misura confinato all’infanzia. Egli sa di essere stato sposato e di essere padre di due figli, ma non sa quasi nulla di concreto sui membri della sua famiglia, che non è in grado di riconoscere né in fotografia né di persona. Il suo sé autobiografico è gravemente compromesso. D’altro canto, al confronto, i ricordi autobiografici di un altro famoso paziente amnesico, Clive Wearing, sono stati risparmiati in misura nettamente superiore. Wearing non ha soltanto un sé nucleare normale, ma anche un robusto sé autobiografico. Un brano tratto da una lettera scrittami da sua moglie, Deborah Wearing, spiega perché io la pensi così:
È in grado di descrivere in modo approssimativo la disposizione della camera di quand’era bambino, sa di aver cantato fin da piccolo nel coro della parrocchia di Erdington, dice di ricordare di essere stato nei rifugi antiaerei durante la guerra e di aver udito il rumore delle bombe su Birmingham. Conosce diversi frammenti di fatti riguardanti la sua infanzia, i suoi genitori e i suoi fratelli, e può raccontare in modo schematico la sua vita adulta: il college a Cambridge, dove era corista; dove ha lavorato; la London Sinfonietta, il Music Department della bbc, la sua camera di direttore d’orchestra, di musicologo e di produttore musicale (e in precedenza di cantante). Ma come le dirà lui stesso, sebbene conosca un vago profilo della sua vita, Clive ha «perso tutti i dettagli».
Negli ultimi tempi, Clive è riuscito a intrattenere conversazioni autentiche e significative meglio di quanto non facesse nei primi dieci anni, quando era pieno di paura e di rabbia. Ha una certa consapevolezza del trascorrere del tempo, giacché parla dei suoi genitori e di suo zio usando il passato (suo zio è morto nel 2003, e dopo che glielo dissi - cosa che lo turbò, poiché erano molto affezionati - non ricordo che abbia mai più parlato di zio Geoff usando il presente). Inoltre, se gli si chiede quanto tempo è passato dalla sua malattia, si arrischierà a dire almeno 20 anni (in realtà sono 25) ; ne ha sempre avuta un’idea approssimativa. Anche in questo caso, non ha alcuna sensazione di saperlo, ma se gli si chiede di fare un’ipotesi, di solito l’azzecca.
Un altro esempio di patologia che può essere attribuita a una compromissione selettiva del sé autobiografico è una condizione nota come anosognosia. In seguito a un danno in una regione dell’emisfero cerebrale destro che comprenda le cortecce somatosensoriali e le cortecce motorie, di solito causato da un ictus, i pazienti presentano, sul lato sinistro, un’evidente paralisi degli arti, soprattutto del braccio; nondimeno, continuano a «dimenticare » di essere paralizzati. Indipendentemente da quante volte sia stato detto loro che non possono muovere il braccio sinistro, quando li si interroga, continuano ad affermare, in tutta sincerità, che esso si muove eccome. Non riescono a integrare l’informazione corrispondente alla paralisi in quel particolare processo in corso di svolgimento che è la storia della loro vita. Relativamente a questi fatti, la loro biografia non è aggiornata, sebbene essi sappiano, per esempio, che hanno avuto un ictus e che sono ricoverati in ospedale. Questo letterale oblio nei confronti di realtà tanto palesi è responsabile dell’apparente indifferenza che essi mostrano per le proprie condizioni di salute e della loro mancanza di motivazione a partecipare alle sedute di riabilitazione di cui avrebbero tanto bisogno.
Devo aggiungere che quando i pazienti subiscono un danno equivalente all’emisfero cerebrale sinistro, non vi è mai anosognosia. In altre parole, il meccanismo grazie al quale aggiorniamo la nòstra biografia circa gli aspetti riguardanti il nostro sistema muscoloscheletrico richiede l’insieme delle cortecce somatosensoriali localizzate dell’emisfero cerebrale destro.
Fenomeni epilettici che si originino in questo stesso sistema possono causare una condizione bizzarra e fortunatamente temporanea: l' asomatognosia. I pazienti conservano il senso del sé e alcuni aspetti della percezione viscerale, ma all’improvviso, e per un breve periodo, non sono in grado di percepire gli aspetti muscoloscheletrici del proprio corpo.
Un ultimo commento sulle patologie della coscienza. Di recente è stato ipotizzato che la corteccia dell’insula sia la base della consapevolezza cosciente degli stati del sentire, e per estensione, della coscienza.20 Ne conseguirebbe che un danno bilaterale a queste cortecce causerebbe un disturbo devastante della coscienza. Da osservazioni dirette, sappiamo che questo non è vero e che i pazienti con un danno insulare bilaterale hanno un sé nucleare normale e una mente cosciente perfettamente attiva.