6
UN’ARCHITETTURA PER LA MEMORIA
IN QUALCHE MODO, DA QUALCHE PARTE
«Chi di noi d’ora innanzi, vedendo partire un treno, non udrà degli spari? »1 chiede ai suoi amici Dick Diver, il protagonista di Tenera è la notte di Scott Fitzgerald, mentre stanno tutti salutando uno di loro, Abe North. A Parigi, una mattina, Diver e compagni hanno appena assistito a qualcosa di inatteso: una giovane donna disperata ha estratto dalla borsetta un piccolo revolver di madreperla e ha sparato al suo amante mentre il treno in partenza usciva fischiando dalla Gare Saint Lazare.
La domanda posta da Diver ci ricorda in modo suggestivo la stupefacente abilità del nostro cervello di apprendere informazioni composite e di riprodurle in seguito con una considerevole fedeltà e da molte prospettive diverse, a prescindere dal fatto che lo si voglia oppure no. D’ora in poi, ogni volta che metteranno piede in una stazione ferroviaria, Diver e i suoi amici udiranno sempre, nella loro mente, spari immaginari, in un’approssimazione vaga ma comunque riconoscibile dei suoni sentiti quella mattina: un involontario tentativo di riprodurre le immagini uditive esperite allora. E poiché il ricordo composito degli eventi può essere rievocato attraverso la rappresentazione di qualsiasi loro parte componente, probabilmente Diver e gli altri udiranno gli spari anche quando qualcuno, in un qualsiasi scenario, si limiterà a menzionare un treno che parte, e non solo quando effettivamente ne vedranno uno uscire da una stazione; e forse li sentiranno anche quando qualcuno accennerà a Abe North (si trovavano là per lui) o alla Gare Saint Lazare (il luogo in cui avviene l’episodio). Questo è anche ciò che accade a chi è stato in una zona di guerra e rivive all’infinito, in tormentosi quanto indesiderati flashback, le scene visive e i suoni della battaglia. La sindrome post-traumatica da stress non è altro che lo sgradevole effetto collaterale di una capacità altrimenti meravigliosa.
Il fatto che, come nel caso di Diver, l’evento da ricordare abbia una sua prominenza emozionale - cioè interferisca con la misura del valore - in genere aiuta. Il cervello memorizzerà scene visive, uditive, tattili, sensazioni, odori e simili, purché esse abbiano un qualche valore e purché al momento fossero sufficientemente permeate di emozione, e le richiamerà al momento giusto. In seguito, l’immagine rievocata potrebbe farsi indistinta. Con il passare del tempo e grazie all’immaginazione di un narratore, il materiale può essere elaborato, fatto a pezzi e ricombinato, dando luogo a un romanzo o a una sceneggiatura. A poco a poco, quelle che erano iniziate come immagini filmiche non verbali possono anche trasformarsi in una descrizione verbale frammentaria, ricordata per le parole del racconto non meno che per i suoi elementi visivi e uditivi.
Consideriamo ora quanto sia meravigliosa questa capacità di ricordare e pensiamo alle risorse che il cervello deve possedere per metterla in campo. In qualche modo, da qualche parte, il cervello - oltre alle immagini percettive riconducibili ai vari domini sensoriali - deve poter archiviare le rispettive configurazioni; in qualche modo, da qualche parte, esso deve anche conservare una via per recuperarle, affinché in qualche modo, da qualche parte, il tentativo di riproduzione funzioni. Nel momento in cui accade tutto questo, e in presenza del dono ulteriore del sé, noi sappiamo che stiamo ricordando qualcosa.
La capacità di manipolare il mondo complesso intorno a noi dipende da questa facoltà di apprendere e ricordare: noi riconosciamo persone e luoghi solo perché registriamo le loro sembianze e poi, al momento opportuno, rievochiamo parte di quelle registrazioni. Anche la nostra abilità di immaginare gli eventi possibili dipende dall’apprendimento e dalla memoria, ed è alla base sia del ragionamento e dell’esplorazione del futuro, sia, più in generale, della creazione di nuove soluzioni per un dato problema. Se vogliamo capire come tutto questo accada, dobbiamo scoprire, nel cervello, i segreti del «come» e localizzare il «dove». Oggi questo è uno dei nodi intricati che le neuroscienze si trovano a dover sciogliere.
L’approccio al problema dell’apprendimento e della memoria dipende dal livello funzionale che decidiamo di studiare. La nostra comprensione di che cosa occorra, a livello di neuroni e piccoli circuiti, affinché il cervello apprenda, è sempre più profonda. A fini pratici, sappiamo come apprendono le sinapsi, e conosciamo perfino alcune delle molecole e dei meccanismi di e-spressione genica implicati nell’apprendimento a livello dei microcircuiti.2 Sappiamo anche che alcune parti specifiche del cervello hanno un ruolo fondamentale nell’apprendimento di tipi diversi di informazione: da un lato vi sono oggetti come facce, luoghi o parole; dall’altro, i movimenti.3 Prima che i meccanismi del « come » e del « dove » possano essere del tutto chiariti, rimane però da rispondere ancora a molte domande. Lo scopo, qui, è di tratteggiare uno schizzo dell’architettura del cervello, che possa dare un ulteriore contributo al chiarimento del problema.
LA NATURA DELLE REGISTRAZIONI MNEMONICHE
Il cervello registra entità - il loro aspetto, i suoni che producono e il modo in cui agiscono - e le preserva per successive rievocazioni. Fa altrettanto anche per gli eventi. Di solito si dà per scontato che il cervello sia un supporto di registrazione passivo - come una pellicola -, sul quale sia possibile mappare fedelmente le caratteristiche di un oggetto, così come vengono analizzate dai rilevatori sensoriali. Se l’occhio è la macchina fotografica innocente e passiva, il cervello è la celluloide vergine, anch’essa passiva. Questa, però, è fantasia bella e buona.
L’organismo (il corpo e il suo cervello) interagisce con gli oggetti, e il cervello reagisce all’interazione. Invece di registrare la struttura di un’entità, il cervello in realtà registra le molteplici conseguenze delle interazioni fra l’organismo e l’entità. Ciò che noi memorizziamo del nostro incontro con un dato oggetto non è soltanto la sua struttura visiva, mappata nelle immagini ottiche della retina. Occorrono infatti anche i seguenti elementi: in primo luogo, le configurazioni sensomotorie associate alla vista dell’oggetto (per esempio i movimenti degli occhi e del collo, o anche del corpo nel suo complesso, se applicabile nel caso specifico); in secondo luogo, la configurazione sensomotoria associata al tatto e alla manipolazione dell’oggetto (se applicabile nel caso specifico); in terzo luogo, la configurazione sensomotoria risultante dall’evocazione di ricordi, acquisiti in precedenza, attinenti all’oggetto; e, infine, le configurazioni sensomotorie legate all’innesco di emozioni e sentimenti relativi all’oggetto.
Quello che normalmente chiamiamo «ricordo dell’oggetto » è il ricordo composito delle attività sensoriali e motorie legate all’interazione fra l’organismo e l'oggetto, nell’arco di un certo periodo di tempo. La gamma delle attività sensomotorie - come pure la loro memorizzazione - varia a seconda del valore dell’oggetto e delle circostanze.
I ricordi di certi oggetti sono governati dal fatto che in passato abbiamo conosciuto oggetti simili o situazioni simili a quelli che stiamo sperimentando. In altre parole, i nostri ricordi sono soggetti a pregiudizio, qui inteso nell’accezione piena del termine: il pregiudizio derivante dalla nostra storia passata e dalle nostre convinzioni. Quello di una memoria perfettamente fedele è un mito, applicabile solo a oggetti banali. L’idea che il cervello possa mai trattenere qualcosa di simile a un « ricordo isolato dell’oggetto» appare insostenibile. Il cervello conserva un ricordo di quanto è accaduto nel corso di un’interazione: una parte importante di quest’ultima comprende il nostro passato e spesso anche il passato della specie biologica e della cultura cui apparteniamo.
Il fatto che la percezione abbia luogo grazie al coinvolgimento, e non mediante una ricettività passiva, è il segreto alla base dell’« effetto proustiano»: la ragione per cui spesso ricordiamo non soltanto gli oggetti isolati, ma anche il loro contesto. D’altra parte, questo fatto è rilevante anche per comprendere il modo in cui la coscienza emerge.
PRIMA LE DISPOSIZIONI, POI LE MAPPE
La caratteristica delle mappe cerebrali è la connessione relativamente trasparente, in esse, fra la cosa rappresentata - forma, movimento, colore, suono - e i contenuti della mappa stessa. All'interno di quest’ultima, la configurazione presenta una qualche inequivocabile corrispondenza con l’oggetto rappresentato. In teoria, un osservatore intelligente, che nel corso dei suoi vagabondaggi scientifici potesse imbattersi nella mappa, indovinerebbe immediatamente quello che si suppone essa rappresenti. Noi sappiamo che questo non è ancora possibile, sebbene le nuove tecniche di neuroimmagine stiano facendo significativi passi avanti in questa direzione. Negli studi su esseri umani condotti con la risonanza magnetica funzionale (fMRI), l’analisi multivariata dimostra la presenza di specifiche configurazioni di attività cerebrale associate a determinati oggetti visti o uditi dal soggetto. In uno studio recente condotto dal nostro gruppo (Meyer et al., 2010, citato nel cap. 3), siamo stati in grado di rilevare, nella corteccia uditiva, configurazioni corrispondenti a quello che i soggetti udivano con l’« orecchio della mente » (senza che in realtà udissero alcun suono). Questi risultati sono una risposta diretta all’interrogativo posto da Dick Diver.
Lo sviluppo biologico delle mappe e la sua diretta conseguenza - le immagini e la mente - rappresentano, nell’evoluzione, una transizione a cui non viene dato sufficiente risalto. Transizione da che cosa? potreste chiedere legittimamente. Da una modalità di rappresentazione neurale che aveva una scarsa connessione esplicita con la cosa rappresentata. Farò un esempio. In primo luogo, immaginiamo che un organismo sia colpito da un oggetto e che, a quel contatto, un gruppo di neuroni risponda scaricando. L’oggetto potrebbe essere appuntito o smussato, grande o piccolo, qualcosa che stava in mano o che era in movimento, fatto di plastica, di acciaio o anche di carne viva. Tutto quello che conta è che esso colpisce l’organismo su una parte della sua superficie, e che a quel punto un gruppo di neuroni risponde alla collisione attivandosi, senza rappresentare effettivamente le proprietà dell’oggetto. Ora immaginiamo un altro gruppo di neuroni che scarichi dopo aver ricevuto un segnale dal primo, e che faccia quindi spostare l’organismo dalla sua posizione stazionaria. Nessuno dei due gruppi di neuroni ha rappresentato dove si trovasse originariamente l’organismo, né dove fosse opportuno che si fermasse, e nessuno dei due gruppi ha rappresentato le proprietà fisiche dell’oggetto. Quello che serviva, qui, era il rilevamento della collisione, un dispositivo di comando, e la capacità di muoversi. Tutto qui. Questi gruppi di neuroni cerebrali non sembrano aver rappresentato mappe, ma piuttosto disposizioni, ossia formule contenenti un know-how e codificanti qualcosa come: se vieni colpito da una parte, muoviti nella direzione opposta per un numero X di secondi, indipendentemente dall’oggetto che ti colpisce o dal luogo in cui ti trovi.
Nel corso dell’evoluzione, per un lungo, lunghissimo tempo, i cervelli hanno operato sulla base di queste disposizioni, e negli ambienti adatti, alcuni degli organismi così attrezzati se la cavavano perfettamente. Il sistema delle disposizioni produsse ottimi risultati e divenne sempre più complicato, realizzando risultati di portata sempre maggiore. Tuttavia, quando si presentò la possibilità delle mappe, gli organismi poterono spingersi oltre le rigide reazioni stereotipe e rispondere invece in base all'informazione più ricca messa ora a disposizione dalla novità. La qualità della gestione migliorò di conseguenza: invece di essere generiche, le risposte si adattarono agli oggetti e alle situazioni, e alla fine acquistarono anche maggior precisione. In seguito, i sistemi disposizionali, che non generavano mappe, unirono le proprie forze a quelli che invece le creavano, e non appena ciò avvenne gli organismi ottennero una flessibilità di gestione ancora maggiore.
L’aspetto affascinante è che il cervello non si liberò del suo sistema già collaudato (le disposizioni) a favore della nuova invenzione (le mappe e le loro immagini). La natura mantenne entrambi i sistemi in funzione e a pieno ritmo: li unì e li fece lavorare in sinergia. Quale conseguenza della combinazione, il cervello si arricchì: in altre parole, divenne il tipo di cervello che gli esseri umani ricevono in dotazione alla nascita.
Ogni volta che percepiamo il mondo, apprendiamo informazioni su di esso, ricordiamo quello che abbiamo appreso e manipoliamo in modo creativo l’informazione, noi umani presentiamo l’esempio più complicato di quella modalità di operazione ibrida e sinergica. Dalle molte specie che ci hanno preceduti, abbiamo ereditato un ricco sistema di disposizioni responsabile del funzionamento dei fondamentali meccanismi di gestione dei processi vitali. Queste reti includono i nuclei che controllano il nostro sistema endocrino e quelli preposti ai meccanismi di ricompensa e punizione, come pure all’innesco e all’esecuzione delle emozioni. In una felice innovazione, le reti disposizionali sono state messe in contatto con molti sistemi di mappe dedicate alla creazione di immagini del mondo interno ed esterno all’organismo. Ecco dunque che i fondamentali meccanismi di gestione dei processi vitali influenzano il funzionamento delle regioni corticali che creano le mappe. Per come la vedo io, d’altra parte, la novità non si ferma qui, e il cervello dei mammiferi si spinse un passo oltre.
Quando decise di creare archivi prodigiosamente vasti di immagini registrate, per risolvere il problema logistico della mancanza di spazio, il cervello degli esseri umani prese a prestito la strategia delle disposizioni. In questo modo, si ritrovò, per così dire, con la botte piena e la moglie ubriaca: riuscì a contenere numerose informazioni in uno spazio limitato, conservando la capacità di richiamarle rapidamente e con una fedeltà notevole. Noi esseri umani, e i mammiferi in genere, non abbiamo mai dovuto fissare su microfilm un gran numero di immagini diverse, conservandole in archivi permanenti: semplicemente, abbiamo memorizzato una formula agile per ricostruirle e abbiamo usato i meccanismi percettivi già esistenti per riassemblarle nel modo migliore possibile. Siamo sempre stati postmoderni.
LA MEMORIA AL LAVORO
Qui sta il problema, allora. Oltre a creare rappresentazioni mappate che danno luogo a immagini percettive, il cervello riesce a mettere a segno un’impresa non meno straordinaria: da un lato, registra nella memoria le mappe sensoriali, dall’altro riproduce, approssimandolo, il loro contenuto originale. Questo processo è noto come recall [lett. « richiamo alla memoria, rievocazione»]. Ricordare una persona o un evento, come pure raccontare una storia, sono operazioni che necessitano del recali; riconoscere oggetti e situazioni intorno a noi richiede, ancora una volta, il recali; lo stesso vale per il pensiero su oggetti con i quali abbiamo interagito in passato e su eventi che abbiamo percepito, e si applica anche, infine, a tutto il processo immaginativo grazie al quale programmiamoli futuro.
Se dobbiamo capire come funziona la memoria, dobbiamo comprendere come - e dove - il cervello registri in forma stabile una mappa. Crea forse un facsimile dell’oggetto da memorizzare, una specie di hard copy archiviata in un file? Oppure riduce l’immagine a un codice - per così dire, la digitalizza? Quale codice? Come? Dove?
Vi è poi un’altra questione essenziale riguardante il dove: durante la rievocazione, dove viene riprodotta la registrazione al fine di recuperare le proprietà essenziali dell’immagine originale? Quando Dick Diver sentirà nuovamente gli spari, dove saranno riprodotti quei suoni nel suo cervello? Quando pensiamo a un amico che abbiamo perduto, o a una casa in cui abbiamo vissuto, noi componiamo un insieme di immagini di quelle entità. Le immagini richiamate alla memoria sono meno intense dell’oggetto reale o di una fotografia; tuttavia possono conservare le proprietà fondamentali dell’originale, al punto che un geniale studioso di neuroscienze cognitive, Steve Kosslyn, è riuscito a stimare le dimensioni relative di un oggetto richiamato alla memoria e ispezionato nella mente.4 Dove vengono ricostruite le immagini, affinché noi possiamo studiarle nella nostra reminiscenza?
Le risposte (« assunti » sarebbe tuttavia un termine migliore) date tradizionalmente a questa domanda si ispirano a una descrizione convenzionale della percezione sensoriale. Grazie ad apposite vie cerebrali, le diverse cortecce sensoriali di ordine inferiore (in larga misura localizzate posteriormente nel cervello) proiettano le varie componenti dell’informazione percettiva alle cosiddette cortecce multimodali (in larga misura localizzate anteriormente), le quali procedono alla loro integrazione. La percezione opererebbe dunque sulla base di una cascata di processori a senso unico: una sequenza che estrarrebbe, un passo dopo l’altro, segnali sempre più raffinati, dapprima nelle cortecce sensoriali di una singola modalità (per esempio quella visiva), e in seguito nelle cortecce multimodali, che ricevono segnali da più di un canale sensoriale (per esempio visivo, uditivo e somatico). In generale, la cascata seguirebbe una direzione caudo-rostrale (ovvero procederebbe dalle regioni posteriori a quelle anteriori) e culminerebbe nella corteccia temporale anteriore e in quella frontale, dove si presume abbiano luogo le rappresentazioni più integrate della percezione multisensoriale della realtà in corso in quel momento.
Il concetto di «cellula della nonna» esprime bene questi assunti. Una cellula della nonna è un neurone localizzato in prossimità del punto più alto della cascata di elaborazione (per esempio, nel lobo temporale anteriore), la cui attività di per se stessa rappresenterebbe in modo completo la nostra nonna nel momento in cui la percepiamo. Queste singole cellule (o piccoli gruppi di queste cellule) sarebbero la sede di una rappresentazione generale completa degli oggetti e degli eventi, durante la percezione. Non solo: esse conserverebbero anche una registrazione di quei contenuti percepiti. La localizzazione delle registrazioni mnemoniche coinciderebbe con quella delle cellule della nonna. Un fatto ancor più impressionante, che risponde direttamente all’interrogativo posto in precedenza, è che - una volta riattivate -le cellule della nonna consentirebbero la riproduzione di quegli stessi contenuti così come sono stati percepiti nella loro interezza. In breve, l’attività di quei neuroni sarebbe responsabile della rievocazione di varie immagini appropriatamente integrate, ivi inclusi il volto di mia nonna o gli spari uditi da Dick Diver alla stazione ferroviaria. Il dove del processo di rievocazione sarebbe dunque quello.
A mio parere, la spiegazione data qui sopra è improbabile. Secondo questa descrizione, un danno che interessi la corteccia frontale e la corteccia temporale anterosuperiore, in altre parole un danno alle regioni anteriori del cervello, dovrebbe precludere sia la percezione normale, sia la normale attività di rievocazione. La prima verrebbe meno perché i neuroni in grado di creare la rappresentazione ben integrata di un’esperienza percettiva coesa non sarebbero più funzionali. Quanto alla seconda, verrebbe meno anch’essa, perché le stesse cellule alla base della percezione integrata sono anche alla base delle registrazioni mnemoniche integrate.
La spiegazione tradizionale - purtroppo per lei - non gode di buona salute: questa previsione infatti non è sostenuta dalla realtà dei risultati prodotti dalle ricerche neuropsicologiche. I punti essenziali di questa realtà discordante sono i seguenti. I pazienti con lesioni localizzate nelle regioni cerebrali anteriori - frontali e temporali - presentano una percezione normale e mostrano soltanto alcuni deficit selettivi nella rievocazione e nel riconoscimento di oggetti ed eventi particolari.
Capita, per esempio, che i pazienti descrivano con gran dovizia di dettagli i contenuti di una fotografia che viene loro mostrata, e che inoltre descrivano la fotografia correttamente come l’immagine di una festa (compleanno, matrimonio), senza tuttavia riuscire a riconoscere che si tratta della loro festa. Un danno alle regioni anteriori del cervello non compromette né la percezione integrata dell’intera scena, né l’interpretazione del suo significato. Non compromette nemmeno la percezione dei numerosi oggetti - persone, sedie, tavoli, torta di compleanno, candeline, vestiti della festa, eccetera - che compongono l’immagine, né il recupero del loro significato. Il danno localizzato anteriormente permette sia la visione integrata sia la visione delle parti. Per compromettere l’accesso alle componenti separabili del ricordo - quelle che corrispondono ai vari oggetti o alle loro caratteristiche, come il colore o il movimento -, occorre una localizzazione del danno completamente diversa. Tale accesso è infatti negato soltanto in seguito a lesioni che interessino settori della corteccia cerebrale situati molto più indietro, nei pressi delle principali regioni sensoriali e motorie.
In conclusione, il danno alle cortecce associative integrative non preclude la percezione integrata, né la rievocazione delle componenti o del significato di un insieme non particolare costituito da più oggetti e caratteristiche. Tali lesioni apportano una sola conseguenza importante e peculiare nel processo di rievocazione: precludono la rievocazione della particolare unicità e della specificità di oggetti e scene. Una determinata festa di compleanno continuerà a essere una festa di compleanno., ma non sarà più la specifica festa di compleanno di qualcuno, associata a un luogo e a una data. Solo un danno che interessi le cortecce sensoriali di ordine inferiore - quelle che creano la mente - o le zone a esse limitrofe preclude effettivamente il richiamo dell’informazione un tempo elaborata da quelle cortecce e registrata nelle regioni adiacenti.
BREVE DIVAGAZIONE SUI TIPI DI MEMORIA
Le possibili distinzioni tra i diversi tipi di memoria riguardano non soltanto la materia che è argomento della rievocazione, ma anche l’insieme delle circostanze in cui quell’argomento è calato, in una particolare situazione di recali. In questa luce, diverse denominazioni tradizionali comunemente applicate a vari tipi di memoria (generica vs. particolare, semantica vs. episodica) non colgono la ricchezza del fenomeno. Per esempio, se qualcuno mi chiede di una particolare casa dove ho vissuto in passato, stimolandomi con un suggerimento verbale o con una fotografia, probabilmente richiamerò alla mente moltissimi elementi legati alle mie esperienze personali in quella casa; questo comporta la ricostruzione di configurazioni sensomotorie di diverso tipo, riconducibili a diverse modalità sensoriali, così che ogni sentimento personale possa essere rivissuto. Se, invece, mi si chiede di rievocare il concetto generale di casa, potrei benissimo richiamare con l’occhio della mente quella stessa casa particolare, e poi accingermi a formulare il concetto generico di casa; in quelle circostanze, tuttavia, la natura della domanda modificherebbe il processo della mia rievocazione. Lo scopo della seconda richiesta, probabilmente, inibirà il ricordo della gran messe di dettagli personali tanto prominenti nel caso precedente: invece di comporre un ricordo personale, mi limiterò a elaborare un insieme di fatti che soddisfino il mio bisogno del momento, che è quello di definire la parola casa.
La distinzione fra il primo e il secondo esempio sta nel grado di complessità insito nel processo del ricordo. Quella complessità può essere misurata in termini di numero e varietà di elementi rievocati in rapporto a un particolare oggetto o evento. In altre parole, quanto più è ampio il contesto sensomotorio che viene rivissuto relativamente a un’entità o a un evento particolari, tanto maggiore è la complessità. Il ricordo di entità ed eventi particolari - precisamente quelli che sono al tempo stesso particolari e personali - richiede contesti di complessità elevata. Qui possiamo cogliere una progressione gerarchica di complessità: le entità e gli eventi particolari-personali richiedono la massima complessità; quelli particolari ma non personali seguono a ruota; infine, quelli non particolari comportano il minimo livello di complessità.
A scopo pratico, è utile affermare che un dato termine viene rievocato a uno solo dei livelli appena specificati: per esempio, «non particolare»; oppure «particolare-personale». Questa distinzione è grossolanamente paragonabile a quella fra memoria semantica/episodica o generica/contestuale.
È anche utile conservare la distinzione fra memoria fattuale e procedurale, perché coglie un fondamentale spartiacque tra gli «oggetti» - entità in stato di quiete, dotate di una certa struttura - e il loro «movimento» nello spazio e nel tempo. Anche qui, però, il confine può diventare insidioso.
In ultima analisi, la validità di queste categorie sta nel fatto che il cervello ne rispetti o meno i confini. Grosso modo, nella rievocazione, il cervello onora le distinzioni fra elaborazione particolare e non; rispetta inoltre la distinzione tra memoria fattuale e procedurale tanto nella creazione di un ricordo, quanto nella sua rievocazione.
UNA POSSIBILE SOLUZIONE DEL PROBLEMA
La riflessione su queste osservazioni mi ha indotto a proporre un modello di architettura neurale mirato a spiegare i processi di rievocazione e riconoscimento.5 Ecco i risultati ottenuti grazie al modello.
Le immagini possono essere esperite sia durante la percezione, sia durante la rievocazione. Sarebbe impossibile archiviare, nel loro formato originale, le mappe alla base di tutte le immagini che sono state oggetto di esperienza. Le cortecce sensoriali di ordine inferiore, per esempio, costruiscono di continuo mappe riguardanti l’ambiente in tempo reale e non hanno risorse per archiviare le mappe scartate. In cervelli come il nostro, però, grazie alle reciproche connessioni fra lo spazio delle disposizioni e quello impegnato nella creazione di mappe, queste ultime possono essere registrate sotto forma di disposizioni, le quali sono quindi anche meccanismi salvaspazio per l’archiviazione delle informazioni. Le disposizioni possono essere usate anche per ricostruire le mappe nello stesso formato in cui erano state originariamente esperite, nelle cortecce sensoriali di ordine inferiore.
Il modello prendeva in considerazione i risultati neuropsicologici descritti in precedenza e postulava che i gruppi cellulari ai massimi livelli delle gerarchie di elaborazione non conservassero rappresentazioni esplicite delle mappe di oggetti ed eventi. Invece, essi conservavano il know-how, ovvero le disposizioni, per procedere -all’occorrenza - alla ricostruzione finale delle rappresentazioni esplicite. In altre parole, stavo usando il semplice sistema delle disposizioni introdotto in precedenza; questa volta però la disposizione, invece di prescrivere un banale movimento, stava ordinando il processo di riattivazione e assemblaggio degli aspetti di percezioni passate, ovunque esse fossero state elaborate e poi localmente registrate. In modo più specifico, le disposizioni agirebbero su molte cortecce sensoriali di livello inferiore originariamente coinvolte nella percezione: lo farebbero per mezzo di connessioni divergenti dal loro sito e dirette alle cortecce sensoriali di livello inferiore. In ultima analisi, il locus in cui le registrazioni mnemoniche verrebbero effettivamente riprodotte non sarebbe molto diverso da quello della percezione originale.
ZONE DI CONVERGENZA-DIVERGENZA
La parte principale del quadro concettuale da me proposto era un’architettura neurale di connessioni corticali con proprietà di segnalazione convergente e divergente rispetto a particolari nodi, che ho chiamato zone di convergenza-divergenza (zcd): esse registravano la coincidenza dell’attività in neuroni localizzati in siti cerebrali diversi, neuroni che erano stati attivati, per esempio, dalla mappatura di un particolare oggetto. Per essere archiviata nella memoria, nessuna parte della mappa complessiva dell’oggetto doveva essere permanentemente ri-rappresentata nelle zcd: occorreva invece che fosse registrata solo la coincidenza dei segnali provenienti dai neuroni connessi alla mappa. Per ricostruire la mappa originale e quindi realizzare la rievocazione, ho proposto il meccanismo della retroattivazione con sincronizzazione. Il termine retroattivazione indicava che il meccanismo necessitava di un processo di « ritorno » per indurre l’attività; il termine sincronizzazione richiamava l’attenzione su un altro requisito, ovvero la necessità di retroattivare le componenti della mappa all’incirca nello stesso intervallo temporale, in modo che quanto aveva luogo simultaneamente (o quasi) a livello di percezione potesse essere ripristinato simultaneamente (o quasi) anche nella rievocazione.
L’altro elemento essenziale del quadro concettuale consisteva nel postulare una divisione del lavoro fra due tipi di sistemi cerebrali: uno che gestiva mappe/immagini e un altro che gestiva disposizioni. Per quanto riguardava le cortecce cerebrali, proposi che lo spazio delle immagini consistesse di diverse isole di cortecce sensoriali di ordine inferiore: per esempio, il gruppo delle cortecce visive che circondano la corteccia visiva primaria (area 17 o VI), l’insieme delle cortecce uditive, quello delle cortecce somatosensoriali, eccetera.
Quanto allo spazio delle disposizioni, esso comprendeva tutte le cortecce associative di ordine superiore delle regioni temporale, parietale e frontale; e inoltre un antico gruppo di sistemi disposizionali localizzato al di sotto della corteccia cerebrale, in strutture quali il prosencefalo basale, i gangli basali, il talamo, l’ipotalamo e il tronco encefalico.
In breve, lo spazio delle immagini è lo spazio in cui si formano le immagini esplicite di tutte le modalità sensoriali: sia quelle che diventano coscienti, sia quelle che rimangono inconsce. Lo spazio delle immagini è localizzato nel grande territorio cerebrale che crea le mappe, formato dall’aggregazione di tutte le cortecce sensoriali di ordine inferiore: regioni corticali localizzate all’inter-
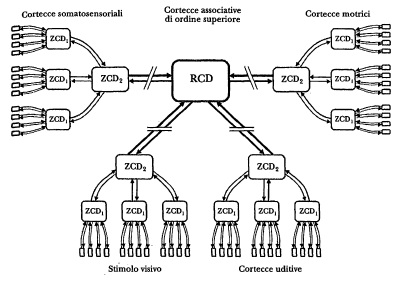
Figura 6.1 Schema dell’architettura di convergènza-divergenza (sono raffigurati quattro livelli gerarchici). Il livello delle cortecce primarie è rappresentato sotto forma di piccoli rettangoli grigi, mentre i tre livelli di convergenza-divergenza (rettangoli più grandi) sono contrassegnati con le sigle zcD2 zcd2 e rcd. Fra i livelli zcd e i livelli RCD (frecce interrotte) possono essere presenti numerose zcd intermedie. Si noti che, in tutta la rete, ogni proiezione in avanti ha una corrispondente proiezione di ritorno (frecce).
no, e nei pressi, del punto in cui segnali visivi, uditivi e di altra natura arrivano al cervello. Esso comprende anche i territori del nucleo del tratto solitario, del nucleo parabrachiale e dei collicoli superiori, che hanno la capacità di formare immagini.
Lo spazio delle disposizioni è quello in cui esse detengono sia la base di conoscenza, sia i dispositivi per la sua ri-costruzione nel processo di rievocazione. È la fonte delle immagini nel processo di immaginazione e ragionamento, ed è anche usato per generare il movimento. È localizzato nelle cortecce cerebrali non altrimenti occupate dallo spazio delle immagini (le cortecce di ordine superiore e parte delle cortecce limbiche) e in numerosi nuclei subcorticali. Quando sono attivati, i circuiti disposizionali inviano segnali ad altri circuiti inducendo la produzione di immagini o azioni.
I contenuti esibiti nello spazio delle immagini sono espliciti, mentre quelli dello spazio disposizionale sono impliciti. Se siamo coscienti, possiamo avere accesso ai contenuti delle immagini, ma non possiamo mai accedere direttamente ai contenuti delle disposizioni. Necessariamente, i contenuti delle disposizioni sono sempre inconsci. Essi esistono in una forma criptata e silente.
Le disposizioni producono diversi risultati. A un livello fondamentale, possono generare azioni di molti tipi e di molti gradi di complessità: la liberazione di un ormone nel sangue circolante; la contrazione della muscolatura dei visceri oppure dei muscoli di un arto o dell’apparato di fonazione. Le disposizioni corticali, però, conservano anche registrazioni di immagini realmente percepite in occasioni precedenti, e partecipano al tentativo di ricostruire un abbozzo di quell’immagine grazie a informazioni memorizzate. Le disposizioni contribuiscono inoltre all’elaborazione delle immagini nel momento in cui sono percepite; per esempio, influenzando il grado di attenzione che viene loro prestato. Noi non siamo mai consapevoli della conoscenza necessaria per eseguire uno qualsiasi di questi compiti, né dei loro passaggi intermedi. Siamo consapevoli soltanto dei risultati, per esempio uno stato di benessere, il battito precipitoso del cuore, il movimento di una mano, il frammento di un suono ricordato, la versione rifinita della percezione di un paesaggio in tempo reale.
I nostri ricordi delle cose, delle proprietà degli oggetti, delle persone e dei luoghi, di eventi e relazioni, di abilità e processi necessari alla regolazione dei processi vitali - insomma tutti i nostri ricordi: quelli ereditati dall’evoluzione e disponibili già alla nascita, e quelli acquisiti in seguito, grazie all’apprendiménto - esistono nel nostro cervello in forma di disposizioni, in attesa di diventare immagini esplicite o azioni. La nostra base di conoscenza è implicita, criptata e non cosciente.
Le disposizioni non sono parole; sono registrazioni astratte di potenzialità. La base per la messa in atto delle parole o dei segni esiste anch’essa sotto forma di disposizioni che poi prendono vita, come immagini e azioni, nella produzione del linguaggio verbale o dei segni. Le regole con cui noi mettiamo insieme parole e segni, la grammatica di un linguaggio, sono anch’esse conservate come disposizioni.
ANCORA SULLE ZONE DI CONVERGENZA-DIVERGENZA
Una zona di
convergenza-divergenza (zcd) è un gruppo di neuroni all’interno del quale
entrano in contatto molti circuiti di feedforward
e
feedback.
Una
zcd
riceve
connessioni feedforward
da aree sensoriali
localizzate a livello «inferiore» nelle catene di elaborazione dei
segnali, le quali iniziano nel punto in cui le afferenze sensoriali
entrano nella corteccia cerebrale. A sua volta, la
zcd
invia
proiezioni feedback
a quelle stesse
aree di origine. Una ZCD invia anche proiezioni feedforward
verso regioni che,
nella catena, sono localizzate al livello di connessione
successivo, ricevendo da esse proiezioni di ritorno.
Le zcd sono microscopiche e si trovano all’interno di regioni di convergenza-divergenza (rcd), che sono invece macroscopiche. Io immagino che il numero delle zcd sia nell’ordine di molte migliaia, e quello delle rcd nell’ordine delle decine. Le zcd sono micronodi; le rcd sono macronodi.
Le rcd sono localizzate, all’interno delle cortecce associative, in aree strategiche verso le quali convergono diverse vie fondamentali. È possibile immaginare le rcd
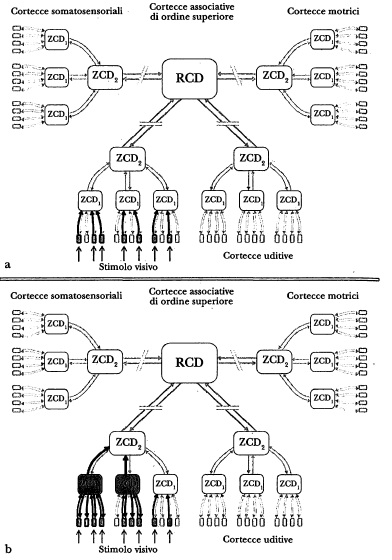
Figura 6.2 Uso dell'architettura di convergenza-divergenza per rievocare ricordi sollecitati da uno specifico stimolo visivo. Negli schemi a e b, un particolare stimolo visivo in entrata (un insieme selettivo di piccoli rettangoli pieni) sollecita l'attività delle zcd dei livelli 1 e 2 (frecce in grassetto e rettangoli pieni). Nello schema evengono attivate specifiche RCD, mentre nello schema d la retroattivazione a partire dalle RCD innesca l’attività di cortecce somatosensoriali, uditive e motrici di ordine inferiore come pure di altre cortecce visive (frecce in grassetto, rettangoli pieni). La retroattivazione genera rappresentazioni nello «spazio delle immagini», ma anche movimenti (insieme selezionato di piccoli rettangoli pieni).
come hub sulla
mappa di una linea aerea. Pensiamo a Chicago, Washington, New York,
Los Angeles, San Francisco, Denver o Atlanta. Questi hub ricevono
gli aeroplani lungo vie di traffico in entrata e li re-inviano
utilizzando le medesime vie. Gli hub - e questo è molto importante
- sono interconnessi anche fra loro, sebbene alcuni siano più
periferici di altri. Infine, alcuni di essi sono più grandi di
altri, il che significa semplicemente che sotto il loro ombrello si
ripara un maggior numero di zcd.
Grazie a studi di neuroanatomia sperimentale, sappiamo che nel cervello dei primati non umani esistono queste configurazioni di connettività.6 Sappiamo inoltre - da recenti studi di neuroimmagine condotti con risonanza magnetica utilizzando diverse tecniche di diffusione - che tali configurazioni esistono anche negli esseri umani.7 Come vedremo nei prossimi capitoli, le rcd sono importanti nella produzione e nell’organizzazione dei contenuti essenziali della mente cosciente, compresi quelli che compongono il sé autobiografico.
Sia le rcd, sia le zcd entrano in essere sotto il controllo genetico. Nel momento in cui l’organismo interagisce con l’ambiente durante lo sviluppo, il rafforzamento o l’indebolimento sinaptico introduce cambiamenti significativi nelle regioni di convergenza e modifica massivamente le zcd. Il rafforzamento sinaptico ha luogo quando le circostanze esterne corrispondono alle esigenze di sopravvivenza dell’organismo.
In breve, il compito che io immagino per le zcd consiste nel ricreare configurazioni separate di attività neurali che durante la percezione si erano verificate in modo all’incirca simultaneo - ovvero coincidenti nella finestra temporale necessaria perché noi prestassimo loro attenzione e ne fossimo coscienti. Per ottenere questo, la zcd indurrà una sequenza di attivazione velocissima - e non avvertita dalla coscienza -, che porterà regioni neurali separate a « connettersi » in un certo ordine.
In questa architettura, il recupero della conoscenza si baserebbe sull’attività - relativamente simultanea e oggetto di attenzione - presente in molte regioni corticali di ordine inferiore, generata nell’arco di diverse reiterazioni di questi cicli di riattivazione. Quelle attività separate sarebbero alla base delle rappresentazioni ricostruite. La conoscenza verrebbe recuperata a un livello dipendente dalla portata dell’attivazione multiregionale. Questa, a sua volta, dipenderebbe dal livello di zcd attivato.8
IL MODELLO ALL’OPERA
Che prove abbiamo del fatto che il modello convergenza-divergenza corrisponda alla realtà? Recentemente, io e il mio collega Kaspar Meyer abbiamo analizzato un gran numero di studi nei campi della percezione, dell’immaginazione e dell’elaborazione «a specchio» e abbiamo considerato i loro risultati dalla prospettiva del modello della convergenza-divergenza:9 molti di essi costituiscono interessanti verifiche del modello in studio. Ecco un esempio.
In una conversazione con un’altra persona, noi ascoltiamo la voce di chi parla e simultaneamente vediamo le sue labbra muoversi. Secondo il modello zcd, poiché un certo movimento delle labbra si verifica ripetutamente in associazione alla componente sonora corrispondente, i due eventi neurali - rispettivamente nelle cortecce visiva e uditiva di ordine inferiore - vengono associati in una zcd condivisa. In futuro, quando ci troveremo di fronte a una sola parte di quella scena (per esempio, se osserveremo uno specifico movimento delle labbra in un videoclip senza audio), la configurazione di attività indotta in tal modo nella corteccia visiva di ordine inferiore attiverà la zcd condivisa, e quest’ultima retroattiverà, nella corteccia uditiva di pari livello, la rappresentazione del suono originariamente associato al movimento delle labbra.
Secondo il modello delle zcd, la lettura labiale, in as-senza di qualsiasi suono, induce l’attività delle cortecce uditive, e le configurazioni di attività così evocate coincidono con quelle indotte durante la percezione di parole effettivamente pronunciate.10 La mappa uditiva del suono diventa parte integrante della rappresentazione del movimento labiale. Il modello zcd spiega come sia possibile, quando si riceve lo stimolo visivo appropriato, udire un suono con l’occhio della mente (o viceversa).
Se qualcuno considerasse cosa di poco conto questa impresa del cervello di sincronizzare elementi visivi e uditivi, dovrebbe pensare al disagio e all’irritazione che si prova quando, nella proiezione di un film, la tecnologia è scadente e le componenti audio e video sono fuori sincronia o, peggio ancora, quando ci tocca guardare un gran film doppiato male. Numerosi altri studi sulla percezione riguardanti altre modalità sensoriali (olfatto, tatto), e anche studi di neuropsicologia condotti su primati non umani, hanno prodotto risultati spiegabili in modo soddisfacente con il modello zcd.11
Altri dati interessanti provengono da alcuni studi sulla creazione di immagini mentali. Come suggerisce il termine stesso, l’immaginazione, intesa come processo, consiste nella rievocazione di immagini e nella loro successiva manipolazione: operazioni di taglio, ampliamento, riordinamento, eccetera. Possiamo chiederci se, quando usiamo la nostra immaginazione, le immagini siano prodotte in forma di «quadri» (visivi, uditivi, eccetera), oppure se essa si basi su descrizioni mentali simili a quelle linguistiche.12 Il modello zcd avalla la prima ipotesi (descrizione in forma di « quadro ») : quando oggetti o eventi sono percepiti direttamente, ma anche quando sono richiamati dall’archivio della memoria, vengono attivate regioni simili. Le immagini costruite durante la percezione sono poi ri-costruite nel processo di immaginazione. Più che repliche, esse sono approssimazioni - tentativi di tornare alla realtà del passato - e quindi non hanno assolutamente la stessa intensità o la stessa accuratezza.
Come indicato in modo inequivocabile da moltissimi studi, l’esecuzione di compiti che implicano la produzione di immagini in modalità visiva o uditiva evoca di solito configurazioni di attività cerebrale considerevolmente sovrapposte a quelle rilevate durante la vera percezione;13 dal canto loro, anche i risultati degli studi su lesioni forniscono prove stringenti della fondatezza del modello zcd in generale e, più in particolare, della descrizione dell’immaginazione come composizione di un quadro. Lesioni cerebrali focali spesso causano deficit simultanei sia a livello della percezione, sia nella produzione di immagini. Un esempio è l’incapacità di percepire e immaginare i colori causata da un danno alla regione occipitotemporale. I pazienti con lesioni focali localizzate in questa regione percepiscono il mondo visivo in bianco e nero: letteralmente, nella gamma dei grigi. Questi soggetti non sono in grado di « immaginare » il colore. Sanno perfettamente che il sangue è rosso, e tuttavia non riescono a immaginare il rosso con l’occhio della mente, non più di quanto possano vederlo quando guardano una. fiche ài quel colore.
Dati provenienti sia da studi di neuroimmagine funzionale, sia da studi di lesioni indicano che, nella corteccia, la rievocazione di oggetti ed eventi si affida, almeno in parte, all’attività che ha luogo sia nei pressi dei siti di ingresso delle afferenze sensoriali, sia nei pressi dei siti di uscita delle efferenze motorie. Di sicuro non è una coincidenza che questi siano gli stessi siti impegnati nella percezione originale di oggetti ed eventi.
Anche la ricerca sui neuroni specchio fornisce le prove del fatto che un’architettura basata sulla convergenza-divergenza offre una spiegazione soddisfacente di certi fenomeni - comportamenti e operazioni mentali -complessi. Nella ricerca sui neuroni specchio (cap. 4) il risultato fondamentale è che la semplice osservazione di un’azione è sufficiente per attivare, nel cervello, le aree motrici corrispondenti.14 Il modelo zcd è ideale per spiegare questa osservazione. Consideriamo che cosa accade quando compiamo un’azione. In primo luogo, un’azione non consiste meramente di una sequenza di movimenti generati dalle regioni motorie del cervello; essa abbraccia rappresentazioni sensoriali che sorgono simultaneamente nelle cortecce somatosensoriale, visiva e uditiva. Secondo il modello zcd, la ripetuta concomitanza delle diverse mappe sensomotorie che descrivono un’azione specifica dà luogo alla ripetizione di segnali convergenti diretti a una particolare zcd. In occasioni successive, quando sarà percepita la stessa azione, per esempio visivamente, l’attività generata nelle cortecce visive attiverà la zcd pertinente. La zcd userà poi retro-proiezioni divergenti verso le cortecce sensoriali di ordine inferiore per riattivare le associazioni relative ad altre modalità, per esempio la somatosensoriale e la uditiva. La zcd segnala inoltre alle cortecce motrici e genera un movimento specchio. Dalla nostra prospettiva, i neuroni specchio sono i neuroni delle zcd implicati nel movimento.15
Secondo il modello zcd, i neuroni specchio, da soli, non potrebbero consentire agli osservatori di cogliere il significato di un’azione. Non sono le zcd a conservare il significato degli oggetti e degli eventi: esse ricostruiscono invece quel significato attraverso una retroattivazione multiregionale sincronizzata che ha luogo in diverse cortecce di ordine inferiore. Poiché è probabile che i neuroni specchio coincidano con le zcd, il significato di un’azione non può essere contenuto in queste cellule soltanto. Occorre invece eseguire una ricostruzione di diverse mappe sensoriali precedentemente associate all’azione: ricostruzione eseguita sotto il controllo delle zcd in cui è stato registrato un legame con quelle mappe originali.16
IL COME E IL DOVE DELLA PERCEZIONE E DELLA RIEVOCAZIONE
La percezione o la rievocazione di moltissimi oggetti ed eventi dipende dall’attività di diverse regioni del cervello coinvolte nella creazione di immagini, e spesso interessa anche regioni legate al movimento. Questa configurazione di attività, così altamente dispersa, ha luogo nello spazio delle immagini. È questa attività, più di quella che ha luogo nei neuroni localizzati al termine delle catene di elaborazione, che ci consente di percepire immagini esplicite di oggetti ed eventi. Da un punto di vista funzionale e anatomico, l’attività al termine delle catene di elaborazione ha luogo nello spazio disposizionale. Quest’ultimo è costituito da zcd e rcd situate nelle cortecce associative, che non producono immagini. Lo spazio disposizionale guida la formazione delle immagini ma non è direttamente coinvolto nella loro presentazione.
In questo senso, lo spazio delle disposizioni contiene le «cellule della nonna», liberamente definite come neuroni la cui attività è correlata alla presenza di un oggetto specifico, ma non come neuroni la cui attività permette, di per se stessa, immagini mentali esplicite di oggetti ed eventi. In effetti i neuroni della corteccia temporale mediale anteriore rispondono, con elevata specificità, a oggetti particolari, tanto nella percezione quanto nella rievocazione: ciò indica che questa regione riceve segnali convergenti.17 La mera attivazione di quei neuroni, d’altra parte, senza la retroattivazione che ne seguirà, non ci consentirebbe di riconoscere la nostra nonna, né di ricordarla. Per far questo, occorre ripristinare una parte sostanziale dell’insieme di mappe esplicite che nel loro complesso ne rappresentano il significato. Come i neuroni specchio, anche i «neuroni della nonna» sono zcd: essi consentono, nelle cortecce sensomotorie di livello inferiore, la retroattivazione multiregionale, con sincronizzazione, di mappe esplicite.
In conclusione, il modello zcd postula due «spazi ce-
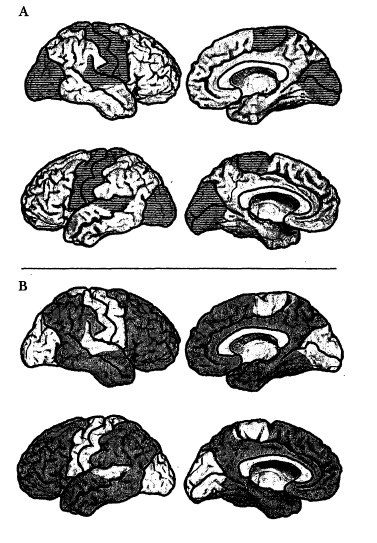
Figura 6.3 Lo spazio delle immagini (mappato) e lo spazio delle disposizioni (non mappato) nella corteccia cerebrale. Nei quattro disegni della tavola A, lo spazio delle immagini, insieme alla corteccia motrice primaria, è rappresentato dalle aree ombreggiate.
Nei quattro disegni della tavola B, le aree ombreggiate rappresentano invece lo spazio delle disposizioni. Le componenti separate dello spazio delle immagini sembrano isole nell’oceano dello spazio delle disposizioni.
rebrali» in un certo qual modo separati. Uno di essi costruisce durante la percezione mappe esplicite di oggetti ed eventi che poi ricostruirà durante la rievocazione. Sia nella percezione, sia nella rievocazione, vi è una evidente corrispondenza fra le proprietà dell’oggetto e la mappa. L’altro spazio, invece, non conserva mappe ma disposizioni, ovvero formule implicite per ricostruire le mappe nello spazio delle immagini.
Lo spazio delle immagini esplicite è costituito dall’insieme delle cortecce sensomotorie di livello inferiore. Quando parlo di « spazio di lavoro » in relazione ai siti in cui vengono assemblate le immagini, penso a esso come al luogo in cui si svolge lo spettacolo di marionette che noi contempliamo nella mente cosciente. Lo spazio delle disposizioni implicite è costituito invece dall’insieme delle cortecce associative. Questo è lo spazio in cui molti inconsapevoli teatranti tirano i fili invisibili delle loro marionette.
I due spazi risalgono a epoche diverse dell’evoluzione del cervello: una in cui le disposizioni erano sufficienti a guidare un comportamento adeguato, l’altra in cui le mappe diedero origine a immagini e a un notevole salto di qualità nel comportamento. Oggi questi due spazi sono integrati senza soluzioni di continuità.