2. IL CERVELLO DI GAGE DISVELATO
Il problema.
All’incirca nella stessa epoca dell’incidente di Gage, i neurologi Paul Broca in Francia e Karl Wernicke in Germania richiamavano l’attenzione degli ambienti medici con i loro studi su pazienti neurologici colpiti da lesioni al cervello. Entrambi, separatamente, suggerivano che i disturbi del linguaggio comparsi in tali pazienti fossero causati da un danno a una ben circoscritta area encefalica (1). La menomazione del linguaggio divenne tecnicamente nota come afasia. Broca e Wernicke pensavano che le lesioni rivelassero, in tal modo, il substrato neurale di due differenti aspetti dell’elaborazione del linguaggio negli individui normali. Le loro tesi erano controverse, e non furono approvate di slancio; ma il mondo ascoltò. Gradualmente, con qualche riluttanza e con molte correzioni, finirono con l’essere accettate. Invece il lavoro di Harlow su Gage e, quanto a questo, anche i commenti di David Ferrier non ricevettero mai la stessa attenzione, e non infiammarono mai in ugual misura l’immaginazione dei colleghi. Molte ragioni possono spiegare il Perchè. Anche se una certa inclinazione filosofica poteva lasciare pensare che il cervello fosse la base della mente, era difficile accettare il punto di vista secondo cui qualcosa vicino all’anima umana (come il giudizio etico) o legato alla cultura (come la condotta sociale) potesse dipendere in misura significativa da una specifica regione cerebrale. Si aggiunga, poi, il fatto che Harlow era un dilettante, rispetto ai professori Broca e Wernicke, e non poteva accampare le convincenti prove che erano necessarie nel suo caso. Questo era soprattutto evidente nella incapacità di indicare una localizzazione precisa del danno al cervello di Gage. Broca era in grado di affermare con sicurezza in quale punto del cervello si trovava il danno che aveva provocato la menomazione del linguaggio (afasia) nei suoi pazienti: di questi egli aveva esaminato e studiato il cervello al tavolo autoptico. Così anche Wernicke, il quale aveva scoperto, dopo la morte di pazienti con menomazioni del linguaggio, che una parte retrostante del lobo temporale sinistro era parzialmente distrutta - e le manifestazioni linguistiche affette erano diverse da quelle identificate da Broca. Harlow non era stato capace di compiere simili osservazioni: egli non solo dovette azzardare una supposta relazione tra la lesione cerebrale di Gage e le alterazioni del suo comportamento, ma dovette in primo luogo ipotizzare una localizzazione della lesione. Così, non era in grado di provare ad alcuno che egli era nel giusto rispetto ad alcunch,.
Ad aggravare l’impaccio di Harlow si aggiungevano le più recenti pubblicazioni di Broca, il quale aveva mostrato che nei suoi pazienti lesioni al lobo frontale sinistro, terza circonvoluzione frontale, erano causa di menomazione del linguaggio (fig. 2.1). L’entrata e l’uscita dell’asta di ferro suggerivano che la lesione al cervello di Gage potesse trovarsi nel lobo frontale sinistro; e però non soffriva di alcuna menomazione del linguaggio, mentre i pazienti di Broca non mostravano alcuna alterazione del carattere. Come potevano esservi risultati così differenti? Con la scarsa conoscenza di neuroanatomia funzionale che era propria di quel tempo, vi fu chi pensò che le lesioni si trovassero approssimativamente nella stessa posizione, e che i differenti risultati non facessero che rivelare la follia di quanti volevano trovare specializzazioni funzionali nel cervello.

Fig. 2.1
B = area di Broca; M = area motoria; W = area di Wernicke. I critici di Harlow sostenevano che la lesione di Gage aveva toccato l'area di Broca, o l'area motoria, o anche tutt'e due e sulla base di tale affermazione attaccavano l'idea di una specializzazione funzionale del cervello umano.
Quando Gage morì, nel 1861, non venne praticata autopsia. Lo stesso Harlow venne a sapere della sua morte solo cinque anni più tardi: l’infuriare della Guerra Civile americana aveva rallentato il propagarsi di notizie di quel genere. Harlow dovette essere rattristato dalla morte di Gage, e tormentato per aver perso l’occasione di studiarne il cervello; tanto che si spinse fino a scrivere alla sorella di Gage per sottoporle una bizzarra richiesta: chiedeva che il corpo fosse esumato in modo che egli potesse averne il cranio e tenerlo a testimonianza del caso.
Ancora una volta, Phineas Gage era l’involontario protagonista di una scena macabra. La sorella e il marito di lei, D. D. Shattuck, insieme con un certo dottor Coon (allora sindaco di San Francisco) e con il medico di famiglia, si rivolsero a un’impresa di pompe funebri, fecero aprire la bara di Gage e ne rimossero il cranio, nonch, la barra di ferro (che, con scrupolo, era stata sepolta assieme al corpo di Gage); poi li mandarono a Harlow. Da allora cranio e barra si trovano a Boston, nel Warren Medical Museum della Harvard Medical School.
Per Harlow, la possibilità di esibire il cranio e la barra era il massimo che egli potesse fare per stabilire che il suo caso non era stato un parto di fantasia, che era davvero esistito un uomo con quella ferita alla testa. Per Hanna Damasio, circa centoventi anni dopo, il cranio di Gage fu il trampolino per un lavoro di investigazione che completò l’opera non finita di Harlow e consente di gettare un ponte tra Gage e la moderna ricerca sulla funzione dei lobi frontali.
La Damasio cominciò con un curioso esercizio: studiare la traiettoria della barra di ferro. Penetrando nella guancia sinistra, diretto verso l’alto, il ferro attraversò, rompendola, la parte posteriore della cavità orbitaria sinistra, posta immediatamente al di sopra. Proseguendo nel suo moto verso l’alto, esso dovette penetrare nella parte frontale dell’encefalo, in prossimità del piano mediano - anche se era difficile indicare esattamente dove. Siccome sembra che fosse inclinato a destra, potrebbe avere colpito dapprima il lato sinistro e poi un po’ sulla destra, mentre muoveva verso l’alto. Il sito iniziale della lesione probabilmente fu la regione orbitaria frontale, direttamente sopra le cavità orbitarie. Nella sua traiettoria, il ferro avrebbe distrutto parte della superficie interna del lobo frontale sinistro, e forse anche di quello destro. Infine, fuoriuscendo, esso avrebbe leso una parte della regione dorsale (o posteriore) del lobo frontale, sicuramente sul lato sinistro e forse anche sul lato destro.
Questa congettura presentava alcune incertezze evidenti. C’era tutta una gamma di potenziali traiettorie che il ferro avrebbe potuto seguire attraverso un cervello “standard”, idealizzato, e non v’era modo di sapere se e quanto tale cervello assomigliasse a quello di Gage. A peggiorare il quadro, si ricordi che anche se la neuroanatomia È gelosa nel tener ferme le relazioni topologiche tra i suoi componenti, vi sono ampie variazioni topografiche da un individuo all’altro, cosicch, ogni cervello umano È molto più diverso dagli altri di quanto lo siano tra loro due autovetture della stessa marca. Si pensi, per chiarire meglio questo punto, alla paradossale uniformità e differenza dei volti umani. Qui c’È un numero invariabile di componenti e una disposizione spaziale invariabile (le relazioni topologiche tra i componenti sono le stesse in tutti i volti); eppure questi volti sono infinitamente diversi, e sono singolarmente distinguibili per via di piccole differenze anatomiche di dimensione, profilo e posizione di quelle parti invariabili e della configurazione (la esatta topografia cambia da volto a volto). Quindi la variabilità individuale rendeva più probabile che la congettura della Damasio fosse erronea.
Hanna Damasio continuò sfruttando la moderna neuroanatomia e i più avanzati metodi di visualizzazione del sistema neurale (2). In particolare, sviluppò e impiegò una nuova tecnica (Brainvox) (3) per ricostruire immagini tridimensionali del cervello di esseri umani viventi, basata sull’elaborazione al calcolatore di dati grezzi ottenuti per scansione ad alta risoluzione del cervello mediante risonanza magnetica. Applicata a soggetti viventi normali o a pazienti neurologici, essa fornisce un’immagine del cervello che non differisce da quella che si vedrebbe sul tavolo autoptico: una meraviglia strana e inquietante. Che cosa avrebbe fatto, il principe Amleto, se avesse potuto contemplare le sue tre libbre di cervello meditabondo ed esitante, e non soltanto il teschio passatogli dal becchino?
Digressione sull’anatomia del sistema nervoso.
Nel capitolo precedente, parlando di frenologia e del legame fra struttura e funzione del cervello, ho accennato all’importanza della neuroanatomia (cioÈ della anatomia del cervello). Torno a insistervi, adesso, Perchè questa È la disciplina fondamentale, per le neuroscienze, dal livello dei singoli, microscopici neuroni (cioÈ le cellule nervose) a quello dei sistemi macroscopici che abbracciano l’intero cervello. In mancanza di una minuziosa conoscenza della sua geografia, a diverse scale, non si può sperare di comprendere i molti livelli di funzioni cerebrali.
Considerando il sistema nervoso nella sua interezza, È facile separarne le divisioni centrali e periferiche. La ricostruzione tridimensionale della figura 2.2 rappresenta il cervello vero e proprio, il componente principale del sistema nervoso centrale. Oltre al cervello, con i suoi due emisferi cerebrali, destro e sinistro, uniti dal corpo calloso (un fitto aggregato di fibre nervose che collegano bidirezionalmente i due emisferi), il sistema nervoso centrale comprende anche il diencefalo (una formazione di nuclei, nascosta sotto gli emisferi, sul piano mediano, comprendente il talamo e l’ipotalamo), il mesencefalo, il midollo allungato, il cervelletto e il midollo spinale.

Fig. 2.2.
Il cervello umano ricostruito in tre dimensioni. L'immagine in alto al centro mostra il cervello visto di fronte; il corpo calloso È nascosto sotto la scissura interemisferica. Le immagini in basso, a sinistra e a destra, mostrano i due emisferi cerebrali separati, con l'indicazione delle principali strutture anatomiche. La copertura a circonvoluzioni degli emisferi È la corteccia cerebrale.
Il sistema nervoso centrale È “neuralmente” connesso con pressoché ogni angolo e recesso del resto del corpo, tramite i nervi: l’insieme di questi costituisce il sistema nervoso periferico. I nervi convogliano gli impulsi dal cervello al corpo e dal corpo al cervello. Come si vedrà meglio nel capitolo 5, tuttavia, cervello e corpo sono anche interconnessi chimicamente, mediante sostanze quali gli ormoni e i peptidi, che sono emessi dall’uno e raggiungono l’altro attraverso il flusso sanguigno.
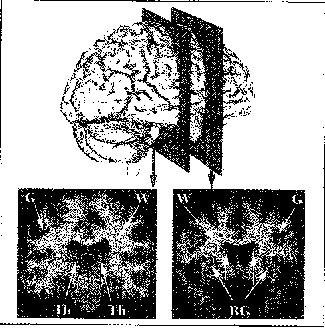
Fig. 2.3.
Due sezioni di un cervello umano vivente ricostruito ottenute mediante risonanza magnetica e mediante la tecnica Brainvox. L'immagine in alto al centro identifica i piani delle sezioni. Si riconosce facilmente la differenza tra sostanza grigia (G) e bianca (W): la prima È ben evidente nella corteccia cerebrale (È il bordo grigio che contorna ogni fessura e ogni protuberanza della sezione) e nei nuclei profondi come i gangli basali (BG) e il talamo (Th).
Una sezione del sistema nervoso centrale permette di comprendere senza difficoltà la differenza tra settori scuri e settori pallidi (fig. 2.3). I primi sono noti come sostanza grigia, anche se in realtà il loro colore di solito È bruno, piuttosto che grigio; i secondi sono noti come sostanza bianca. La sostanza grigia corrisponde in larga misura a raccolte di corpi cellulari (di cellule nervose), mentre la sostanza bianca corrisponde in larga misura ad assoni, o fibre nervose, uscenti dai corpi cellulari nella sostanza grigia.
Quest’ultima si presenta in due varietà. In una i neuroni sono disposti a strati, come in una torta, e formano una "corteccia": ne sono esempi la corteccia cerebrale, che copre gli emisferi cerebrali, e la corteccia cerebellare, che avvolge il cervelletto. Nella seconda varietà di sostanza grigia, i neuroni non sono stratificati, ma sono invece organizzati come noci di acagiù in una ciotola: essi formano un "nucleo". Vi sono nuclei grandi come il caudato, il putamen e il "globus pallidus", celati nella profondità di ciascun emisfero, o l’amigdala, nascosta all’interno di ciascun lobo temporale; vi sono ampie raccolte di nuclei più piccoli, come quelli che formano il talamo; e poi piccoli nuclei singoli, come il "locus niger" o il "nucleus coe-ruleus", nel midollo allungato.
La struttura alla quale le neuroscienze hanno dedicato il massimo impegno di studio È la corteccia cerebrale: la si può immaginare come un rivestimento che copre tutte le superfici del cervello, comprese quelle situate nella profondità delle fenditure note come scissure e solchi, che danno al cervello il suo caratteristico aspetto ripiegato (fig. 2.2). Questo rivestimento È spesso circa 3 millimetri ed È formato da diversi strati, paralleli tra loro e alla superficie del cervello (fig. 2.4). Tutta la sostanza grigia che si trova sotto la corteccia (i nuclei, grandi o piccoli, e la corteccia cerebellare) È denominata subcorticale. Si chiama neocorteccia (o neopallio) la parte più giovane - in senso evolutivo - della corteccia cerebrale. La maggior parte della corteccia evolutivamente più antica È nota come corteccia limbica (si veda più avanti). Nel corso dell’esposizione, farò riferimento di solito o alla corteccia cerebrale (intendendo la neocorteccia) o alla corteccia limbica e ad alcune sue parti specifiche. La figura 2.5 rappresenta una mappa (di uso frequente) della corteccia cerebrale, in cui sono indicate le diverse aree citoarchitettoniche - cioÈ sono distinte le regioni aventi architettura cellulare differente. E’ nota come mappa di Brodmann, e le sue aree sono contraddistinte da numeri. Una divisione del sistema nervoso centrale alla quale farò più volte riferimento È quella, sia corticale sia subcorticale, che È nota come sistema limbico. (Il termine È un po’ un’etichetta generale per un certo numero di strutture evolutivamente antiche: molti neuroscienziati sono riluttanti a usarlo, ma sovente si rivela utile). Le strutture principali che lo costituiscono sono il giro del cingolo, nella corteccia cerebrale, e l’amigdala e il prosencefalo basale, due collezioni di nuclei.

Fig. 2.4.
A = schema dell'architettura cellulare della corteccia cerebrale, con la sua tipica struttura a strati; B = schema dell'architettura cellulare di un nucleo.
Il tessuto nervoso (o neurale) È costituito da cellule nervose (neuroni) sostenute da cellule gliali (gliociti). I neuroni sono le cellule essenziali per l’attività cerebrale e in ogni cervello ve ne sono a miliardi; sono organizzati in circuiti locali i quali, a loro volta, costituiscono le regioni corticali (se sono disposti in strati) o i nuclei (se sono riuniti in insiemi non stratificati). Infine, i nuclei e le regioni corticali sono interconnessi in modo da formare sistemi, e sistemi di sistemi, a livelli via via più alti di complessità. In termini di scala, tutti i neuroni e i circuiti locali sono microscopici, mentre le regioni corticali, i nuclei e i sistemi sono macroscopici.

Fig. 2.5.
Mappa delle principali aree del cervello individuate da Brodmann nei suoi studi dell'architettura cellulare (citoarchitettura). La figura serve solo come riferimento anatomico, non È una mappa frenologica Né una mappa dell'epoca con la rappresentazione delle funzioni cerebrali. Alcune aree sono troppo piccole per poter essere riportate qui, oppure si trovano nascoste nelle profondità di solchi e scissure. In alto, l'aspetto esterno dell'emisfero sinistro; in basso, l'aspetto interno.
Il neurone È costituito da tre importanti componenti: il corpo cellulare; il prolungamento principale in uscita, una fibra nervosa detta assone o cilindrasse; i prolungamenti in entrata, le fibre nervose dette dendriti (fig. 2.6). I neuroni sono collegati tra loro in circuiti in cui si riconoscono gli equivalenti dei fili conduttori (le fibre degli assoni) e dei connettori (le sinapsi, cioè i punti in cui gli assoni sono in contatto con i dendriti di altri neuroni).

Fig. 2.6.
Un neurone con i suoi componenti principali: corpo cellulare, asso-ne, dendriti.
Quando i neuroni diventano attivi (“scaricano” o “sono eccitati”, nel gergo delle neuroscienze), dal corpo cellulare si propaga lungo l’assone una variazione di potenziale elettrico, o potenziale di azione. Arrivato a una sinapsi, questo innesca l’emissione di particolari sostanze chimiche note come neurotrasmettitori (un esempio È il glutammato). A loro volta, i neurotrasmettitori operano sui recettori. In un neurone eccitatorio, l’interazione in cooperazione con molti altri neuroni le cui sinapsi sono adiacenti, e che possono liberare oppure no i propri neurotrasmettitori, determina se il prossimo neurone sarà eccitato oppure no, cioÈ se esso produrrà il proprio potenziale di azione, con la conseguente emissione dei propri neurotrasmettitori, e così di seguito.
Le sinapsi possono essere forti o deboli. La forza di una sinapsi determina se e quanto facilmente gli impulsi continueranno a propagarsi verso il neurone successivo. In generale, in un neurone eccita-torio, una sinapsi forte facilita il cammino degli impulsi, mentre una sinapsi debole lo ostacola o lo arresta (4).
E’ opportuno qui fare un accenno alla natura della connettività tra neuroni. Non pochi scienziati, quando si trovano di fronte alla complessità delle connessioni tra neuroni, disperano di riuscire, un giorno, a comprendere il cervello. Alcuni preferiscono nascondersi dietro la considerazione che ogni cosa si connette con ogni altra cosa e che il comportamento, con buona probabilità, emerge da tale volente o nolente connettività in modi che la neuroanatomia non saprà mai rivelare. Ma costoro, per fortuna, si sbagliano. E infatti, si osservi che ogni neurone forma, in media, un migliaio di sinapsi (anche se ve ne sono con 5000 o 6000 sinapsi). Può sembrare un numero grande, ma quando si pensa che vi sono più di 10 miliardi di neuroni e più di 10000 miliardi di sinapsi, ci si rende conto che ogni singolo neurone È moderatamente connesso. Si prendano - a caso, o seguendo le proprie preferenze anatomiche - alcuni neuroni della corteccia o dei nuclei, e si troverà che ognuno di essi “parla” con pochi altri. In effetti, molti neuroni parlano solo con altri neuroni che non sono molto distanti nell’àmbito di circuiti relativamente locali di nuclei e regioni corticali; altri, seppure dotati di assoni che si protendono per diversi millimetri (o addirittura per centimetri) nel cervello, entreranno in contatto soltanto con un numero relativamente piccolo di altri neuroni. La conseguenza principale di tale disposizione È che qualsiasi cosa i neuroni facciano dipende dal gruppo di neuroni che li circonda e di cui fanno parte; che qualsiasi cosa i sistemi facciano dipende dal modo in cui i gruppi influenzano altri gruppi, in un’architettura di gruppi interconnessi; che qualsiasi contributo di ciascun gruppo alla funzione del sistema cui appartiene dipende dal posto che occupa in tale sistema. In altre parole, la specializzazione del cervello cui si È fatto cenno nella “Digressione sulla frenologia” del capitolo 1 È una conseguenza del posto occupato da raggruppamenti di neuroni connessi in modo rado all’interno di un sistema a più ampia scala.
In breve: il cervello È un supersistema di sistemi. Ognuno di questi È composto da una elaborata interconnessione di regioni corticali e nuclei subcorticali, piccoli ma macroscopici, che sono fatti di microscopici circuiti locali; questi ultimi sono fatti di neuroni, i quali sono tutti interconnessi mediante sinapsi. (Non di rado si trovano i termini “circuiti” e “rete” impiegati come sinonimi di “sistema”; È bene precisare se si fa riferimento alla scala micro - o macroscopica. Nel presente libro, salvo indicazione diversa, i sistemi sono macroscopici e i circuiti sono microscopici).
Livelli di architettura neurale:
- Neuroni
- Circuiti locali
- Nuclei subcorticali
- Regioni corticali
- Sistemi
- Sistemi di sistemi.
La soluzione
Dal momento che per Phineas Gage non si poteva ricorrere alla scansione, Hanna Damasio pensò a un approccio indiretto al suo cervello (5). Per far questo, si procurò l’aiuto di Albert Galaburda, un neurologo della Harvard Medical School, il quale andò al War-ren Medical Museum e fotografò con molta cura il cranio di Gage da differenti angolazioni; inoltre misurò le distanze tra le zone ossee colpite dalla barra e alcuni punti di riferimento standard sul cranio. L’analisi di queste fotografie, combinata con le descrizioni della ferita, consentì di restringere la gamma delle possibili traiettorie della barra. Inoltre le fotografie consentirono alla Damasio e a un suo collega neurologo, Thomas Grabowski, di ricreare il cranio di Gage in coordinate tridimensionali, e di ricavarne le più probabili coordinate di un cervello che si adattasse a tale cranio. A questo punto subentrò un altro collaboratore, Randall Frank, un ingegnere, il cui contributo consentì di compiere una simulazione in un calcolatore di grande potenza. Essi riprodussero una barra di ferro tridimensionale con le stesse misure di quella di Gage, e la “infilzarono” in un cervello - prossimo per forma e dimensioni a quello di Gage - secondo le possibili traiettorie che essa avrebbe potuto seguire nell’incidente. Le figure 2.7 e 2.8 mostrano i risultati.

Fig. 2.7.
Fotografia del cranio di Gage, fatta nel 1992.
Adesso siamo in grado di confermare l’asserzione di David Ferrier, secondo la quale, anche se una parte non trascurabile di cervello andò perduta, la barra non toccò le regioni cerebrali necessarie per il linguaggio o per la funzione motoria. (Le aree intatte di entrambi gli emisferi comprendevano le cortecce motoria e premotoria, come pure l’opercolo frontale, sul lato sinistro noto come area di Broca).
Si può affermare con sicurezza che il danno fu più esteso sul lato sinistro che su quello destro, e nei settori anteriori anzich, in quelli posteriori della regione frontale nel suo insieme. Il danno compromise le cortecce prefrontali per lo più nelle superfici ventrale e interna di entrambi gli emisferi, risparmiando invece in larga parte il fronte laterale, o esterno, delle cortecce prefrontali.

Fig. 2.8.
In alto, ricostruzione del cervello e del cranio di Gage con la possibile traiettoria della barra di ferro. In basso, vista dall'interno dei due emisferi cerebrali, per mostrare come la barra lese le strutture dei lobi frontali su entrambi i lati.
In Gage fu sicuramente danneggiata parte di una regione che dalle nostre più recenti osservazioni si rivela critica per il processo di decisione in condizioni normali, cioÈ la regione prefrontale ventrome-diana. (Nella terminologia neuroanatomica, la regione orbitaria È nota anche come regione ventromediana del lobo frontale, e questo È il termine che io userò nel seguito. Tale regione È, per così dire, il “bassoventre” del lobo frontale; “mediana” indica prossimità al piano mediano o alla superficie interna di una struttura). La ricostruzione rivelò che in Gage non furono danneggiate regioni ritenute vitali per altri aspetti della funzione neuropsicologica. Per esempio, rimasero intatte le cortecce sulla faccia laterale del lobo frontale; un danno arrecato a esse distrugge la capacità di controllare l’attenzione, di eseguire calcoli e di passare opportunamente, quando occorre, da uno stimolo all’altro.
Questa moderna ricerca permise di trarre alcune conclusioni. La Damasio e i suoi colleghi poterono affermare con qualche fondamento che era stato un danno selettivo alle cortecce prefrontali del cervello di Phineas Gage quello che aveva compromesso la sua capacità di comportarsi nel rispetto delle regole sociali a lui note in precedenza, di decidere la linea di condotta capace di risultare alla fine la più vantaggiosa per la sua sopravvivenza, di pianificare il proprio futuro. Quel che mancava, adesso, era la conoscenza del possibile modo di operare della mente di Gage quando egli si comportava nel modo rovinoso che effettivamente mostrò dopo l’incidente; ma per questo occorreva esaminare, e studiare le controparti moderne di Gage.