La evolución del desarrollo celular
Lewis Wolpert
LEWIS WOLPERT, miembro de la Royal Society, es catedrático de biología aplicada a la medicina en el Colegio Universitario de Londres. Su interés principal es la morfogénesis embrionaria. Ha escrito, entre otras obras, Triumph of the Embryo (1991).
El desarrollo celular es el mayor logro de la evolución natural. Esto puede sonar algo presuntuoso si comparamos la aparente sencillez de la célula con la complejidad de organismos como los seres humanos, dotados de cerebros extraordinarios. Pero, en términos evolutivos, estos organismos pluricelulares no aparecieron hasta que las células hubieron evolucionado. Es más, creo que, una vez surgida la célula eucariota (dotada de movimiento, un núcleo y orgánulos como las mitocondrias) a partir de bacterias más sencillas, la evolución de estructuras complejas como el cerebro fue, en comparación, relativamente simple.
La evolución como modificación del desarrollo
Los procesos del desarrollo son fundamentales para la evolución de los seres pluricelulares. La evolución avanza mediante modificaciones en el programa de desarrollo del embrión, que a su vez determinan las características del organismo adulto. Esta modificación se debe a cambios en los genes que controlan el desarrollo. En palabras del genetista molecular francés François Jacob, la evolución hace chapuzas con los embriones, utilizando sus piezas para construir nuevas estructuras. Este proceder salta a la vista cuando observamos los embriones jóvenes de los vertebrados, todos asombrosamente parecidos entre sí en cierta etapa del desarrollo (la llamada fase filotípica) a partir de la cual se diferencian (figura 1). La evolución chapucea con el plan corporal básico.

Figura 1. Los embriones de los vertebrados convergen en una fase filotípica común y luego divergen. Arriba, el huevo fecundado; en el centro, la fase filotípica; abajo, el estado adulto.
El desarrollo de las extremidades es un buen ejemplo. Mientras que la forma de las extremidades se ha mantenido estable en muchos vertebrados terrestres, su desarrollo ha sido «chapuceado» para producir alas en las aves y los murciélagos, patas en los caballos y manos en los seres humanos. Es más, las propias extremidades evolucionaron a partir de las aletas de los peces. Otros ejemplos espectaculares los proporcionan los huesos del oído medio de los mamíferos, que en nuestros antepasados reptilianos formaban parte de la articulación de la mandíbula, y el sistema genitourinario, en el que el riñón primitivo se modificó para formar parte del sistema reproductor y evolucionó un nuevo riñón.
Otra característica fundamental de la evolución ha sido la modificación del crecimiento. Esto puede observarse en la evolución de la pezuña del caballo, cuyos dedos laterales crecen más despacio que los centrales. A medida que el caballo crece, los dedos laterales se hacen mucho más pequeños que los centrales y el dedo medio acaba soportando la mayor parte del peso. Los cambios en la estructura craneal de los vertebrados también se deber básicamente al proceso de crecimiento (figura 2). Tal como señaló D’Arcy Thompson en su libro Sobre el crecimiento y la forma, el crecimiento diferencial puede explicar las diferencias básicas en la forma de los cráneos.
La evolución puede contemplarse como la modificación del desarrollo. Así pues, es esencial comprender tanto el desarrollo mismo como su evolución.
Desarrollo
La relación entre las células y los organismos pluricelulares se establece, salvo raras excepciones, a través del desarrollo de una única célula: el huevo. (Una excepción son los mixomicetes celulares, cuyos cuerpos fructíferos se desarrollan por agregación de gran número de amebas.) Todos los animales y plantas que vemos a nuestro alrededor se han desarrollado a partir de una única célula, el huevo fecundado, cuyo desarrollo se caracteriza por una sucesión de divisiones celulares que originan una masa de entre un centenar y un millar de células más pequeñas. Esta masa de células primordiales se diferencia en muchos tipos celulares distintos, como las células musculares y las cartilaginosas. Estos tipos celulares producen a su vez estructuras bien definidas, como las extremidades, en un proceso denominado formación de patrones. Las diferencias entre los vertebrados, sean aves, seres humanos, hipopótamos o chimpancés, se deben fundamentalmente a esta formación de patrones, a la organización espacial de las células, y no a diferencias significativas entre los tipos célulares que los componen. Por poner un ejemplo, los seres humanos no tenemos ningún tipo celular que no esté presente en los chimpancés. Un tercer proceso de desarrollo es el cambio en la disposición de las capas celulares. En la gastrulación, las células superficiales que formarán el conducto digestivo y el esqueleto se desplazan hacia el interior del embrión. La organización del embrión se completa cuando éste es aún muy pequeño (las extremidades humanas adquieren su estructura antes de que el embrión supere el centímetro de longitud) y las estructuras grandes son producto del crecimiento en etapas posteriores. Toda esta estructuración implica un número relativamente pequeño de células, y las interacciones celulares son de carácter local. Una vez se ha establecido el plan corporal básico, el desarrollo de órganos específicos, como los ojos o las extremidades, es relativamente autónomo.
El resultado del desarrollo embrionario está determinado por el comportamiento de las células individuales, y este comportamiento celular está dirigido por el ADN. No obstante, la actuación del material genético es más bien pasiva. El ADN sólo contiene códigos para la fabricación de proteínas, que a su vez determinan cómo se comporta la célula. Las enzimas que catalizan las reacciones celulares básicas son proteínas, que también forman parte de la composición de elementos estructurales como el citoesqueleto y la membrana. Las células se caracterizan por las proteínas específicas que contienen (como la queratina de la piel o la hemoglobina de los glóbulos rojos) distintas de las proteínas genéricas que mantienen las funciones celulares básicas. Otro tipo importante de proteína son los factores de transcripción que controlan la activación y desactivación de los genes y, con ello, los tipos de proteínas que fabrica la célula. Así pues, el tipo y el estado de la célula están determinados por la activación o desactivación de ciertos genes y, en consecuencia, por las proteínas presentes. El ADN del embrión no contiene una descripción del organismo adulto sino, más bien, un conjunto de instrucciones para fabricarlo mediante una secuencia de síntesis de proteínas. La papiroflexia es un buen modelo para este proceso. En este caso, las instrucciones corresponderían a la síntesis de proteínas específicas a medida que procede el desarrollo.

Figura 2. Las formas de los cráneos de un ser humano (arriba) y un chimpancé (abajo) reflejan básicamente cambios de crecimiento diferencial.
A continuación describiré algunos de los mecanismos de estructuración básicos implicados en este proceso, pues para entender la evolución del desarrollo embrionario es necesario conocer sus características principales.
Uno de los mecanismos de formación de patrones se basa en la información posicional (figura 3). La idea básica es que la localización de las células no se especifica mediante un sistema de coordenadas, sino que éstas adquieren identidades posicionales (o direcciones) respecto de ciertas fronteras. Una vez fijada la identidad posicional, la célula puede interpretarla mediante un comportamiento específico que depende de su pasado. Este cambio de comportamiento puede originar un tipo celular diferenciado o una modificación de la forma o la adherencia de la célula. Otra interpretación posible es la especificación de un programa de crecimiento. Una característica importante de este mecanismo de estructuración es que no hay una relación simple entre la especificación posicional y la estructura final, sino que ambas están disociadas. Es más, una de las características más atractivas de la información posicional es que las mismas identidades posicionales pueden especificar un gran número de estructuras, lo cual está apoyado por numerosas observaciones. El carácter de cada estructura depende de la manera en que las células interpretan su identidad posicional.

Figura 3. La información posicional puede utilizarse para especificar patrones espaciales. Cada posición queda especificada por la concentración local de un «morfógeno» que es leída por las células.
La información posicional puede proporcionar un concepto unificador para la comprensión del desarrollo regulado de estructuras muy diversas. En principio, las únicas interacciones intercelulares implicadas son las que especifican la posición. Es más, las mismas señales e identidades posicionales pueden utilizarse para generar estructuras bastante diferentes, cuya variedad reflejaría tanto la constitución genética de la célula como la historia de su desarrollo.
La especificación del diseño corporal
Las primeras etapas del desarrollo implican la especificación del diseño corporal, es decir, la determinación de los ejes principales (anteroposterior y dorsoventral) y de regiones concretas como, por ejemplo, los lugares donde se forman la cabeza y las extremidades. Esto define un bosquejo del futuro organismo y, en los animales segmentados, establece un patrón periódico. En los vertebrados, la especificación del diseño corporal hace que todos los embriones pasen por una etapa filotípica común (figura 1) en la que es muy difícil distinguir entre embriones de especies diferentes y a partir de la cual divergen los desarrollos posteriores. También se aprecia una divergencia previa a la etapa filotípica cuando retrocedemos hasta el huevo inicial. Las primeras etapas del desarrollo de mamíferos y ranas son bastante diferentes, pero ambos tipos de embriones convergen luego en la fase filotípica. Esta cuestión evolutiva es tan importante como la conservación de la etapa filotípica.
Los detalles de las primeras etapas del desarrollo no deben preocupamos, de manera que nos centraremos en la especificación del diseño corporal que conduce a la fase filotípica. Empezaremos por los insectos, que también pasan por una fase filotípica, porque son los organismos de los que se tiene más información acerca del control genético de las primeras etapas del desarrollo embrionario. La mosca del vinagre, del género Drosophila, es el organismo mejor estudiado ahora mismo, y la comprensión de las primeras etapas de su desarrollo ha tenido una gran repercusión en el estudio de otros sistemas.
Los ejes anteroposterior y dorsoventral de Drosophila se especifican ya en la fase de huevo. Así, existe un gen llamado bicoid que codifica un factor de transcripción cuyo ARN mensajero se localiza en el extremo anterior del huevo. Tras la fecundación se establece un gradiente de proteína bicoide a lo largo del eje anteroposterior (figura 4) que, al parecer, proporciona información posicional a las células del embrión (aunque en estas primeras etapas no hay paredes celulares entre los núcleos). De esta forma las células pueden «conocer» su distancia al extremo anterior a partir de la concentración de proteína bicoide. A ciertas concentraciones críticas se activan determinados genes que, a su vez, codifican factores de transcripción capaces de activar o inhibir otros genes. Se establece así una sucesión de activaciones de genes que determina la regionalización de varios factores de transcripción de proteínas. Estos factores activan pares de genes con siete «bandas» de actividad de unos pocos núcleos de amplitud, lo cual define catorce parasegmentos precursores de los segmentos corporales de la larva y la mosca adulta. Cada una de las bandas de actividad de estos genes queda especificada por la concentración local de los factores de transcripción que actúan en la región controlada por los genes.

Figura 4. La información posicional en el huevo de Drosophila se especifica inicialmente mediante el gradiente de concentración de la proteína bicoide, que se sintetiza en el extremo anterior del huevo (A) y se difunde hacia el extremo posterior del mismo.
En una etapa posterior, equivalente a la fase filotípica, cada segmento adquiere una identidad específica en virtud de los genes activos en su interior. Estos genes codifican factores de transcripción que confieren identidad posicional a los segmentos, y pertenecen al denominado «complejo génico homeótico». Los genes de este complejo se localizan en un mismo cromosoma y tienen algunas características interesantes. En primer lugar, se llaman así porque sus mutaciones pueden producir homeosis, es decir, transformaciones de una estructura en otra. Por ejemplo, hay mutaciones de estos genes que hacen que se desarrolle una pata allí donde debería formarse una antena, o que un segmento adquiera la identidad de otro adyacente, lo que da lugar a una mosca de cuatro alas. En segundo lugar, el orden de los genes en el cromosoma coincide con el orden en el que se expresan a lo largo del eje anteroposterior. Así, los genes situados en posición anterior y posterior en el complejo se expresan en el extremo anterior y posterior del embrión, respectivamente. En tercer lugar, todos los genes contienen una pequeña región similar conocida como homeobox, o «caja homeótica», que codifica una parte de la proteína que se liga al ADN.
La presencia de esta caja homeótica es importante porque se han encontrado regiones parecidas en casi todos los animales. Los vertebrados poseen cuatro complejos génicos homeobox, cuya expresión en el eje anteroposterior sigue el mismo orden observado en la mosca. En general, se piensa que los genes de las cajas homeóticas registran identidades posicionales a lo largo del eje principal del cuerpo. Es como si la evolución hubiera encontrado una manera satisfactoria de registrar las diferencias posicionales y la hubiera usado una y otra vez. Todos estos complejos génicos tienen su origen en algún antepasado primitivo, y los cuatro complejos de los vertebrados deben haber surgido mediante duplicaciones de esos mismos genes en un antepasado posterior.
Por desgracia, en el caso de los vertebrados aún no se sabe bien cómo se consigue que la información genética del complejo homeobox se traduzca en la posición correcta, pero es seguro que hay implicado algún mecanismo de señalización intercelular.
Alas y extremidades
El desarrollo de las extremidades de los vertebrados no sólo es importante por sí mismo, sino que también nos proporciona un modelo útil. Existe una gran similitud entre los procesos de estructuración de las extremidades y los de especificación del plan corporal.
Las extremidades del pollo brotan de los costados, después de lo cual se especifican sucesivamente los elementos esqueléticos: primero el húmero, luego el radio y el cúbito, luego la muñeca y finalmente los dedos. Los dedos son bastante peculiares y, por convención, se designan con los números 2, 3 y 4, siendo 2 el anterior y 4 el posterior. (En nuestras manos, el dedo meñique es el posterior.) Centrémonos ahora en la estructuración de los dedos.
Al parecer, la región polarizadora, situada en el borde posterior del esbozo de la extremidad, es la fuente de alguna señal. Si se injerta una región polarizadora en el margen anterior de otra extremidad se obtiene un miembro que es su imagen especular (figuras 5a y 5b), de manera que en vez del patrón normal 2 3 4 tenemos el patrón 4 3 2 2 3 4. La región polarizadora establece un nuevo conjunto de identidades posicionales en la región anterior del esbozo, posiblemente una señal decreciente que forma el dedo 4 a un umbral de activación alto y el dedo 3 a un umbral de activación bajo. Estudios recientes muestran que el gen sonic hedgehog se expresa en la región polarizadora, y la proteína que codifica es una buena candidata para esta señal posicional.
Esta misma pauta de señalización a lo largo del eje anteroposterior se encuentra en los esbozos de los miembros anteriores y posteriores de todos los amniotas, lo que explica que una región polarizadora de ratón pueda especificar imágenes especulares de miembros en el pollo. Es más, el gen hedgehog podría estar implicado en la señalización posicional de otras regiones del embrión, como puede ser la especificación del eje principal del cuerpo y la disposición de las neuronas en la médula espinal. Esto sugiere que una misma señal de posición puede actuar en diferentes etapas del desarrollo.
También existe un parecido extraordinario entre el desarrollo de las extremidades de los vertebrados y el del ala de Drosophila. Aquí el gen hedgehog, que vuelve a estar implicado, se activa en una banda estrecha a lo largo del eje del ala en desarrollo. Parece ser que este gen especifica un «morfógeno» cuya difusión originaría la estructura de las regiones anterior y posterior, de manera que, cuando se expresa en una posición inusual, actúa como si fuera el injerto de una región polarizadora y especifica un nuevo conjunto de estructuras alares.
Conservación de los mecanismos del desarrollo
Esta breve disquisición sobre los procesos del desarrollo muestra que a lo largo de la evolución se han conservado mecanismos comunes a sistemas tan diversos como los insectos y los vertebrados. Los genes que especifican el estado celular y las moléculas señalizadoras pueden ser similares en organismos distintos. Buen ejemplo de ello son los complejos homeobox y la familia de moléculas señalizadoras hedgehog, que se utilizan una y otra vez. Un ejemplo reciente y sorprendente es el descubrimiento de que genes similares están implicados en el desarrollo de los ojos de insectos y vertebrados.
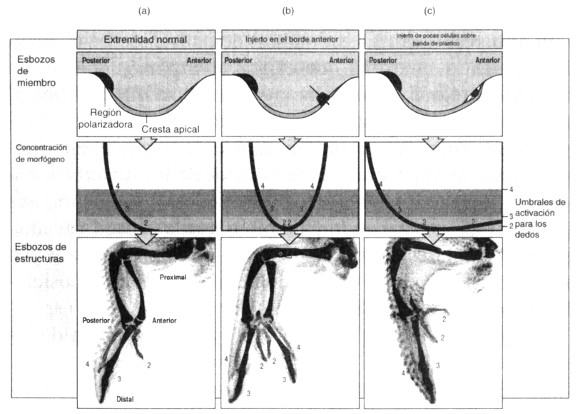
Figura 5. (a) La región polarizadora de la extremidad del pollo determina la identidad de los dedos. El mecanismo podría basarse en un morfógeno difusivo producido por la región polarizadora. (b) Si se injerta una región polarizadora en la región anterior, aparece una extremidad lateralmente invertida, (c) Si se injertan sólo unas pocas células polarizadoras la señal se atenúa y sólo se produce una duplicación parcial.
Esta conservación de los mecanismos del desarrollo embrionario no debe eclipsar su innegable diversidad, sobre todo en las primeras etapas. Los animales y las plantas utilizan una gran diversidad de mecanismos para establecer los ejes en el huevo. El eje dorsoventral del embrión de los anfibios lo especifica el punto de entrada del espermatozoide, mientras que el óvulo de los mamíferos tiene simetría esférica y sus ejes no se especifican hasta mucho más tarde. En la ascidia, el futuro músculo se especifica en gran parte por procesos citoplasmáticos y no mediante interacciones celulares.
Sin embargo, en cierto sentido, la evolución ha sido conservadora, incluso indolente, a la hora de inventar mecanismos de desarrollo. Una vez se «descubrieron» mecanismos capaces de generar organismos complejos, éstos se utilizaron una y otra vez. Así, los mecanismos morfogenéticos basados en la información posicional y la generación de estructuras periódicas producen una variedad enorme, casi infinita, de patrones y estructuras, sobre todo cuando se acoplan con cambios de forma y crecimiento. No hay nada especial en el desarrollo del cerebro humano, que en su conjunto hace uso de mecanismos heredados de nuestros antepasados primitivos.
El origen del embrión
La célula es un sistema maravillosamente complejo, tanto que la interacción entre sus componentes es superior en muchos órdenes de magnitud a la que existe entre las células de un embrión. En este sentido, podemos decir que una célula es mucho más compleja que un embrión. El desarrollo embrionario no es más que el resultado de actividades celulares modificadas de forma coordinada.
La aparición de la célula hace unos 3000 millones de años puede considerarse como la «gran explosión» de la evolución biológica, si bien fue un proceso que requirió mucho tiempo. En comparación, el origen del desarrollo embrionario puede verse como una «pequeña explosión», posterior a la evolución de la célula eucariota. ¿Qué se requería, ni términos generales, para que evolucionaran los procesos del desarrollo? ¿Cómo se originaron el huevo, la generación de patrones y los cambios de forma? Puesto que el desarrollo embrionario requiere la formación de un organismo pluricelular a partir de una única célula, el origen del huevo es un problema fundamental.
La evolución del desarrollo está ligada al origen de los seres pluricelulares, los metazoos. Se cree que los eucariotas unicelulares existen desde hace unos 1400 millones de años, mientras que los restos más antiguos de metazoos tienen unos 800 millones de años de edad. Esto sugiere que la transición de los eucariotas unicelulares a la pluricelularidad fue difícil y requirió cientos de millones de años. No obstante, está claro que el registro fósil de aquellos delicados ancestros primitivos es muy fragmentario e incompleto. Lo que yo creo es que, una vez hubo células eucariotas con capacidad para replicarse y desplazarse, los elementos básicos que requiere el desarrollo ya estaban disponibles, por lo que la transición a la pluricelularidad pudo ser relativamente sencilla. Aun así, tuvieron que transcurrir unos cuantos cientos de millones de años hasta la explosión del Cámbrico, momento a partir del cual se encuentran fósiles de animales reconocibles como tales.
Los procesos básicos (diferenciación, organización espacial, cambio de forma y crecimiento) ya estaban presentes en la célula eucariota. El ciclo celular (es decir, la secuencia de eventos que determina el crecimiento y división de la célula) ilustra todos estos procesos.
En el ciclo celular tiene lugar una secuencia de actividades génicas que puede considerarse un precursor simple de un programa de desarrollo. A medida que la célula crece se activan y desactivan genes en las distintas fases del ciclo celular, las cuales pueden verse como estados diferenciados. También se producen movimientos intracelulares y una reordenación espacial que asegura el reparto equitativo de los cromosomas entre las dos células hijas. Es más, a medida que las sustancias se acumulan, aparecen umbrales de activación que marcan la transición de una fase a la siguiente. El mecanismo de división celular, la mitosis, permitía también la diferenciación de las células hijas a través de la distribución desigual de los componentes celulares durante la división celular. Esta característica ya está presente en las levaduras, cuyas células hijas pueden tener «sexos» diferentes.
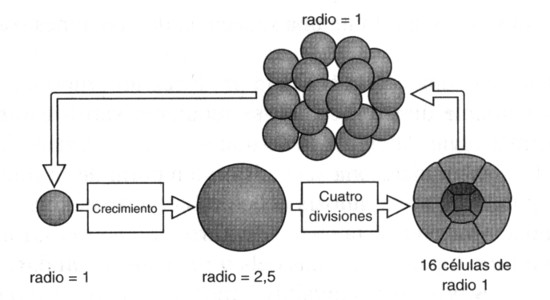
Figura 6. El origen del embrión a partir del agrandamiento y posterior división múltiple de una célula.
A continuación expondré un camino hipotético por el que la célula eucariota pudo llevar al desarrollo embrionario. Un requisito central es que cada etapa tenga una ventaja selectiva y haya continuidad entre las etapas. Los saltos grandes (monstruos prometedores) no están permitidos. Aun así, reconozco que mi hipótesis no es mucho mejor que alguno de Los cuentos de así fue de Rudyard Kipling, del estilo de «Así fue como el leopardo consiguió sus manchas» o «Así fue como el dromedario consiguió su joroba».
Hay dos teorías principales sobre el origen de los metazoos y, por ende, sobre el origen del desarrollo embrionario. La primera especula que la pluricelularidad se derivó de la agregación de dos o más células para formar una colonia; la segunda, en cambio, propone un crecimiento celular con multiplicación nuclear y establecimiento posterior de fronteras celulares. Mi propuesta es una variante de esta última teoría. Lo que pretendo ofrecer es un mecanismo que permita explicar la identidad genética de todas las células del embrión y, sobre todo, el origen del huevo. El mecanismo se basa en el crecimiento de una sola célula más allá de su tamaño normal, seguido de una secuencia de divisiones o escisiones.
Para que una célula solitaria produzca un grupo pluricelular mediante división celular se requieren varios cambios. En primer lugar, la célula tiene que superar su tamaño habitual, lo que requiere una suspensión temporal de la mitosis. En segundo lugar, la mitosis debe desbloquearse en la célula agrandada para que ésta pueda dividirse varias veces. En tercer y último lugar, las células deben permanecer unidas.
Los dos primeros requisitos implican la modificación del ciclo celular, y es relativamente sencillo sugerir procesos basados en mecanismos de control observables en las células actuales. Se ha visto que, durante la mitosis, el tamaño de las levaduras se incrementa en un medio de cultivo rico y disminuye en un medio pobre. Es muy posible, por lo tanto, que el momento de la transición dependa de señales ambientales. Por ejemplo, una célula que creciera en un medio rico podría aumentar de tamaño y, tras ser trasplantada a un medio pobre, experimentar dos divisiones seguidas sin crecimiento. Así ocurre en el protozoo Chlamydomonas, cuyas células se dividen en cuatro o incluso ocho en cada ciclo celular cuando la tasa de crecimiento es rápida.
Así pues, podemos imaginar una célula que incremente su diámetro dos veces y media y luego se divida cuatro veces para producir dieciséis células hijas (figura 6). Supongamos además que se cumple el tercer requisito, es decir, que las células se mantengan juntas y formen una esfera hueca. Esto último requiere divisiones celulares orientadas o uniones intercelulares en la superficie exterior, lo cual no parece muy difícil de conseguir.
El resultado final de estos cambios sería un incremento del tamaño celular en un medio rico, conducente a una esfera pluricelular en un medio pobre. Esto podría ser una ventaja selectiva si las células fueran ciliadas, pues la esfera podría desplazarse más deprisa en busca de ambientes más ricos. Tenemos así por primera vez una selección positiva del estado pluricelular en un medio pobre, y dicho estado habría sido inducido por el ambiente.
Otro «cuento de así fue» para el origen de la pluricelularidad y del huevo se basa en la muerte celular y el canibalismo. Imaginemos una mutación tal que, cuando una célula se divide, las dos células hijas permanecen unidas. Esto podría haber proporcionado a este pequeño organismo una ventaja selectiva muy importante sobre los organismos unicelulares en momentos de penuria: mientras estos últimos morían de hambre, nuestro antepasado bicelular podría haber sobrevivido a los malos tiempos utilizando una célula como alimento de la otra. La célula superviviente sería el origen del huevo, pues las células somáticas de todos los organismos pluricelulares que se desarrollan a partir de un huevo se sacrifican para la supervivencia de éste. En muchos organismos hay células especiales cuya única función es asistir el desarrollo del huevo. Así pues, nuestros orígenes podrían radicar en el canibalismo o en el suicidio altruista.
A continuación propondré un mecanismo basado en un proceso similar hasta cierto punto al efecto Baldwin, formulado a principios de siglo y generalizado por el embriólogo británico Conrad Waddington en lo que él llamaba «asimilación genética». En esencia, este mecanismo implica la inclusión en el programa de desarrollo de un efecto de origen ambiental a través del reemplazo de la señal ambiental por una señal embrionaria. Mediante el efecto Baldwin podemos imaginar mutaciones tales que, en cualquier tipo de medio, las células superen su tamaño normal y luego se dividan varias veces. Una señal ambiental habría sido asimilada por los genes y, en cierto sentido, se habría incorporado al programa. La ventaja selectiva de la pluricelularidad podría ser una mayor velocidad de natación, una mejora en la alimentación o la compartición de metabolitos.
Sólo falta un paso más para que la evolución dé lugar al embrión: que las células individuales se separen y el programa empiece de nuevo. Esto podría haber sucedido cuando las células hubieran crecido demasiado para mantenerse unidas. Luego cada célula podría haber vuelto a pasar por el mismo programa, y habría surgido el desarrollo embrionario. Esta etapa simple, aunque crucial, pudo requerir mucho tiempo. En este embrión primitivo, al que llamaremos Blastaea, cada célula se comporta como un huevo y no hay organización espacial. Los orígenes de la estructuración y la organización espacial, que llevan a la especialización celular, podrían encontrarse en el momento en que los mecanismos de los ciclos celulares alterados generaran una esfera hueca, formada por una única capa de células.
La clave del desarrollo es la generación de diferencias entre las células. Sólo así puede estructurarse el organismo para que se produzcan cambios de forma organizados y se diferencien tipos celulares en lugares específicos. El desarrollo del alga Volvox ilustra algunos de estos principios. La división de su huevo produce células somáticas y germinales distribuidas en forma de esfera hueca. La línea germinal se especifica mediante escisiones desiguales de las que surgen células hijas de tamaños diferentes, las mayores de las cuales dan lugar a células germinales.
¿Cómo evolucionaron estos mecanismos basados en la información posicional? Supongamos que los Blastaea primitivos habitaran en el fondo del mar (figura 7). No es absurdo suponer que las células en contacto con el sustrato se diferenciaran en cuanto a metabolismo o receptores superficiales. Con el tiempo, esta señal ambiental podría haber inducido una sucesión de actividades a partir del punto de contacto, que afectaría a las células locales y produciría señales capaces de afectar a células distantes. Por ejemplo, las células capaces de unirse al punto de contacto podrían tener una ventaja selectiva. Otra ventaja podría derivarse de la invaginación de las células en el punto de contacto. Fuese cual fuese la ventaja, una señal ambiental produciría un cambio local en el organismo, que se haría más complejo con el tiempo. Esto podría acarrear la supresión del crecimiento de las células adyacentes y, en consecuencia, la restricción de la reproducción en el extremo posterior del embrión. De esta forma se especificaría un eje embrionario. El cambio gradual de la señal a lo largo de este eje podría haber originado un patrón basado en la información posicional. De hecho, hay ejemplos actuales de estos procesos, como son la polarización del huevo del alga Fucus por la luz o la determinación del eje dorsoventral de los anfibios a partir del lugar de entrada del espermatozoide en el óvulo.

Figura 7. Origen hipotético de un eje en el punto de contacto de Blastaea con el sustrato.
Volviendo al efecto Baldwin, podemos conjeturar cómo se incorporó al programa de desarrollo la especificación de un eje ambientalmente inducido. Todos los elementos para dicha especificación ya estaban presentes; lo único que se requería era reemplazar la señal ambiental por otra de origen celular. Esto podría conseguirse mediante una diferenciación local del citoplasma del huevo que pudiera definir células germinales, un fenómeno común en los animales actuales.
Otros cambios evolutivos posteriores que implicarían duplicación de genes podrían originar nuevos tipos celulares y los mecanismos de generación del patrón segmentado. Los mecanismos para establecer las identidades posicionales mediante cajas homeóticas habrían evolucionado pronto, y la combinación de todos estos procesos proporcionaría un medio potente para generar una gran variedad de organismos pluricelulares.
Gastrulación
La especificación del plan corporal implica la generación de un patrón espacial, pero el desarrollo también implica cambios importantes en la forma del embrión, el más espectacular de los cuales es la gastrulación. Durante este proceso, las células superficiales del embrión se mueven hacia el interior. Este desplazamiento es necesario porque las células del mesodermo y del endodermo (que originarán el esqueleto y el intestino) se especifican inicialmente en una capa externa del embrión. Este tipo de movimiento es bastante complejo, y la mejor manera de visualizarlo es representar el embrión primitivo como un globo esférico. Si se coloca el dedo en el punto que correspondería al ano y se empuja hasta perforar el otro extremo, se define el lugar de la boca, y el tubo resultante corresponde al intestino. La gastrulación tiene lugar en la mayoría de animales, y su persistencia evolutiva quizá se deba a que es mucho más sencillo estructurar tejidos en un espacio bidimensional (como la superficie de una esfera) y crear después una tercera dimensión extendiendo el crecimiento de la capa hacia el interior. Estos cambios de forma siempre requieren fuerzas generadas por ciertas células cuya disposición depende de la generación de un patrón espacial previo.

Figura 8. Un origen hipotético de la gastrulación. Blastaea forma un intestino primitivo para capturar mejor los organismos de los que se nutre. Esto se parece a la gastrulación que da lugar al intestino en los erizos de mar.
La evolución de la gastrulación hizo posible obtener embriones complejos con dos y hasta tres capas celulares. El biólogo decimonónico alemán Ernst Haeckel sugirió que la gastrulación se originó en un organismo primitivo, al que llamó Gastraea. Haeckel conjeturó que sus predecesores, los Blastaea, se establecieron en el fondo marino. Un pequeño plegamiento hacia dentro habría constituido una ventaja selectiva si hubiera permitido al Blastaea recoger e ingerir alimento de forma más eficaz (figura 8). Esta invaginación habría aumentado de tamaño y se habría convertido en un sistema digestivo primitivo. Tan sólo faltaba que el extremo de la invaginación se uniera a la capa del otro extremo (como sucede en los erizos de mar) para obtener un verdadero intestino. Es posible que el repliegue de las células originara el mesodermo. Es de destacar la cantidad de animales actuales, como el erizo de mar, cuya gastrulación se aproxima mucho a este proceso primitivo hipotético.
¿Qué presiones selectivas se ejercen sobre el embrión en desarrollo? En comparación con el organismo adulto o la larva, el embrión es un ser bastante privilegiado. No tiene que buscar comida o pareja y, en consecuencia, no entra en la competencia por el nicho ecológico. Su función primordial es desarrollarse de manera fiable, y esta fiabilidad es la principal propiedad objeto de selección. Esto no excluye en absoluto la selección de aspectos del desarrollo relacionados con la reproducción y los ciclos vitales, como son la cantidad de vitelo presente, la velocidad del desarrollo y la evolución de formas larvales.
Si la presión selectiva principal recae sobre la fiabilidad del desarrollo, entonces es posible que las variaciones que no afecten a este concepto no sean objeto de selección negativa. Así, la expresión de genes en células equivocadas, la secreción de moléculas a destiempo o las invaginaciones transitorias podrán mantenerse siempre que no interfieran en el resultado final. No está probado que tales procesos sean demasiado costosos, lo que los haría objeto de selección negativa. En cambio, es asombroso lo que cuesta mantenerse vivo: las bombas de sodio y calcio consumen ellas solas un 25% de la energía disponible, mientras que la síntesis y degradación de proteínas consume otro 50%. Esta ausencia de selección negativa ofrece al embrión la posibilidad de explorar nuevos caminos morfogenéticos. Puesto que su principal condicionante selectivo es la fiabilidad de su desarrollo, los embriones, como los estudiantes universitarios veleidosos, pueden llevar una vida disoluta siempre que, al final, aprueben los «exámenes».
Consideremos, por ejemplo, los Blastaea o Gastrea primitivos antes de la evolución del endodermo. Imaginemos que un reducido número de células se desplaza hacia dentro debido a una mutación aleatoria. Estas células podrían persistir durante muchas generaciones y podrían diferenciarse de muchas maneras diferentes. Algunas de estas formas serían eliminadas por la selección, mientras que otras podrían generar, por ejemplo, músculos internos. La presión selectiva principal recae sobre el adulto. Las modificaciones de la señalización molecular y la expresión génica también posibilitan la emergencia de combinaciones útiles.
Otra consecuencia importante de la posición privilegiada del embrión es que hace muy probable que distintas vías morfogenéticas conduzcan a un mismo resultado. No hay una forma de gastrulación que sea «mejor» que otra; es el resultado final lo que importa. De hecho, existen muchas variantes, como en las vías que conducen a los embriones primitivos de dos capas de algunos invertebrados.
Conclusiones
No es difícil imaginar caminos hipotéticos que conduzcan de la célula eucariota a los organismos pluricelulares. También se puede conjeturar cómo se diferenciaron los huevos de las células corporales y cómo evolucionaron los ejes corporales y los patrones espaciales. La evolución de estos procesos habría involucrado inicialmente señales ambientales, que luego se habrían independizado en virtud del efecto Baldwin. La duplicación génica y la segmentación, así como la generación de patrones basada en la identidad posicional y la división celular asimétrica, abrieron vías para la divergencia de nuevas pautas de desarrollo.
A partir de estos mecanismos básicos se generó una gran diversidad de organismos. Es concebible que estos mecanismos morfogenéticos se seleccionaran porque generaban variedad con relativa facilidad. Por ejemplo, un mecanismo basado en la información posicional disocia la generación de diferencias de la manera en que se desarrollan las células. El mismo conjunto de identidades posicionales puede generar multitud de patrones diferentes. Todo ello, junto con la posición evolutivamente privilegiada del embrión, pudo facilitar el desarrollo de organismos cada vez más complejos.
BIBLIOGRAFÍA
Akam, M., P. Holland y G. Wray (eds), «The evolution of developmental mechanisms», Development Supplement (1994).
Lawrence, P., The Making of a Fly, Blackwell, Oxford, 1992.
Maynard Smith, J. y E. Szathmáry, The Major Transitions in Evolution, W. H. Freeman/Spektrum, Oxford, New York, Heidelberg, 1995.
Raff, R. A., The Shape of Life: Genes Development and the Evolution of Animal Form, University of Chicago Press, Chicago, 1996.
Waddington, C. H., The Strategy of the Genes, Alien & Unwin, Londres, 1957.
Wolpert, L., The Triumph of the Embryo, Oxford University Press, Oxford, 1992.