Homo homini lupus («El hombre es un lobo para el hombre») es un antiguo proverbio romano que popularizó Thomas Hobbes. Aun cuando su tesis básica impregna buena parte del derecho, la economía y las ciencias políticas, el proverbio encierra dos grandes errores. En primer lugar, no hace justicia a los cánidos, que son unos de los animales más gregarios y cooperativos del planeta (Schleidt y Shalter, 2003). Y lo que es aún peor, el proverbio niega la naturaleza intrínsecamente social de nuestra propia especie.
La teoría del contrato social, y con ella la civilización occidental, parece imbuida de la suposición de que somos criaturas asocíales, incluso malvadas, en lugar del zoon politikon que Aristóteles vio en nosotros. Hobbes rechazó explícitamente la idea aristotélica cuando propuso que al principio nuestros antepasados eran autónomos y combativos y establecieron la vida comunitaria sólo cuando el coste de los conflictos se volvió insoportable. Según Hobbes, la vida social nunca llegó a nosotros de forma natural. La consideraba un paso que dimos a regañadientes y «sólo mediante un pacto, lo cual es artificial» (Hobbes, 1991 [1651], pág. 120). En fecha más reciente, Rawls (1972) propuso una versión más moderada de la misma idea al añadir que el paso de la humanidad hacia la socialidad dependía de que se dieran condiciones de justicia, es decir, de la posibilidad de una cooperación mutuamente beneficiosa entre iguales.
Estas ideas sobre el origen de la sociedad bien ordenada siguen estando muy extendidas, incluso pese a la suposición subyacente de que es insostenible, a la luz de lo que sabemos acerca de la evolución de nuestra especie, una decisión racional por criaturas intrínsecamente asociales. Hobbes y Rawls crean la ilusión de una sociedad humana que responde a un acuerdo voluntario con reglas autoimpuestas consentidas por agentes libres e iguales. Sin embargo, nunca hubo un momento en el que devinimos sociales: descendemos de ancestros altamente sociales —un largo linaje de monos y simios— y siempre hemos vivido en grupo. Nunca ha existido la gente libre e igual. Los humanos empezamos siendo —si es que se puede distinguir un punto de partida— seres interdependientes, unidos y desiguales. Procedemos de un largo linaje de animales jerárquicos para los que la vida en grupo no es una opción, sino una estrategia de supervivencia. Cualquier zoólogo clasificaría nuestra especie como obligatoriamente gregaria.
Tener compañeros ofrece unas ventajas inmensas a la hora de localizar alimento y evitar a los predadores (Wrangham, 1980; Van Schaik, 1983). En tanto que los individuos con una orientación grupal dejan más descendencia que aquellos con tendencias menos sociales (por ejemplo, Silk y otros, 2003), la socialidad se ha vuelto cada vez más arraigada en la biología y psicología de los primates. Por tanto, de haberse tomado cualquier decisión de crear sociedades, el mérito debería atribuirse a la madre naturaleza y no a nosotros mismos.
No pretendemos con esto rechazar el valor heurístico de la «postura original» de Rawls como una forma de hacernos reflexionar sobre el tipo de sociedad en la que nos gustaría vivir. Su postura original se refiere a una «situación puramente hipotética caracterizada para llegar a determinadas concepciones de justicia» (Rawls, 1972, pág. 12). Pero incluso si no aceptamos la postura original al pie de la letra, y sólo la adoptamos por el bien de la argumentación, sigue distrayendo del argumento más pertinente que deberíamos estar persiguiendo, que es cómo hemos llegado a ser lo que somos. ¿Qué partes de la naturaleza humana nos han conducido hasta aquí, y cómo han determinado esas partes la evolución? Al abordar un pasado real y no hipotético, estas cuestiones nos acercan a la verdad, que es que somos profundamente sociales.
Un buen ejemplo de la naturaleza plenamente social de nuestra especie es que, después de la pena de muerte, el castigo más extremo que podemos concebir es el confinamiento en solitario. Y esto es así, sin duda, porque no hemos nacido para solitarios. Nuestros cuerpos y nuestras mentes no están diseñados para vivir en ausencia de otros. Nos deprimimos sin apoyo social: nuestra salud se déteriora. En un experimento reciente, los voluntarios sanos que se expusieron deliberadamente a un virus del resfriado y la gripe enfermaban con más facilidad si tenían pocos amigos y familiares a su alrededor (Cohén y otros, 1997). Aunque las mujeres comprenden de forma natural la primacía de la conexión con los demás —quizá porque durante 180 millones de años las hembras mamíferas con tendencias que priman el cuidado de los otros se han reproducido más que las que no tenían tales tendencias—, lo mismo se puede aplicar a los hombres. En la sociedad moderna no hay una forma más eficaz de que los hombres amplíen su horizonte de vida que casarse y permanecer casados: incrementa su esperanza de vida más allá de los 65 años entre un 65 y un 90 % (Taylor, 2002).
Nuestra naturaleza social es tan evidente que no sería necesario insistir en este aspecto de no ser por su notoria ausencia de explicaciones sobre el origen de nuestra especie en las disciplinas del derecho, la economía y la ciencia política. La tendencia occidental a ver las emociones como signo de debilidad y los vínculos sociales como algo caótico ha hecho que los teóricos recurran a la cognición como la guía predilecta del comportamiento humano. Celebramos la racionalidad. Y lo hacemos pese a que las investigaciones psicológicas sugieren la primacía del afecto: es decir, que el comportamiento humano deriva ante todo de juicios emocionales rápidos y automatizados, y sólo secundariamente de procesos conscientes más lentos (por ejemplo, Zajonc, 1980, 1984; Bargh y Chartrand, 1999).
Por desgracia, el énfasis en la autonomía individual y la racionalidad, y el correspondiente descuido de las emociones y el afecto, no se limita a las humanidades y las ciencias sociales. También en la biología evolutiva hay quien ha adoptado la idea de que somos una especie autoinventada. Se ha avivado un debate paralelo que enfrenta la razón y la emoción con respecto al origen de la moralidad, un rasgo distintivo de la sociedad humana. Una corriente de pensamiento considera que la moralidad es una innovación cultural conseguida únicamente por nuestra especie. Esta corriente no considera las tendencias morales como algo perteneciente a la naturaleza humana. Sostiene que nuestros ancestros se volvieron morales por elección. La segunda corriente, por el contrario, considera que la moralidad es prolongación directa de los instintos sociales que compartimos con otros animales. Según esta última, ni la moralidad nos pertenece en exclusiva, ni es una decisión consciente adoptada en un momento temporal concreto: es el producto de la evolución social.
El primer punto de vista presupone que, en el fondo, no somos verdaderamente morales. Considera que la moralidad es un revestimiento cultural, una fina capa que oculta una naturaleza egoísta y brutal. Hasta fecha reciente, éste era el enfoque dominante de la moralidad en la biología evolutiva, así como entre los divulgadores científicos que han popularizado este campo. Utilizaré la expresión «teoría de la capa» para designar estas ideas, cuyo origen se debe a Thomas Henry Huxley (aunque obviamente se remontan mucho más allá en la filosofía y la religión occidentales, hasta llegar a la noción del pecado original). Tras abordar estas ideas, examino el punto de vista, bastante diferente, de Charles Darwin sobre una moralidad fruto de la evolución, inspirado por el Siglo de las Luces escocés. Analizo a continuación las ideas de Mencio y Westermarck, que coinciden con las de Darwin.
Dadas estas opiniones contrarias sobre la continuidad frente a la discontinuidad respecto de otros animales, me basaré en un estudio anterior (De Waal, 1996) en el que presto una atención especial a la conducta de los primates no humanos para explicar por qué creo que los fundamentos de la moralidad son antiguos desde el punto de vista evolutivo.
LA TEORÍA DE LA CAPA
En 1893, ante un numeroso público en Oxford, Inglaterra, Huxley reconcilió públicamente su sombría visión del mundo natural con la amabilidad que encontraba ocasionalmente en la sociedad humana. Huxley era plenamente consciente de que las leyes del mundo físico son inalterables. Sin embargo, creía que era posible mitigar y modificar su impacto en la existencia humana si la gente mantenía a la naturaleza bajo control. Así, Huxley comparó a los humanos con un jardinero que tiene muchas dificultades para impedir que las malas hierbas se apoderen de su jardín. Consideraba que la ética humana constituye una victoria sobre un proceso evolutivo ingobernable y terriblemente desagradable (Huxley, 1989 [1894]).
Se trataba de una postura asombrosa por dos razones. En primer lugar, ponía freno deliberadamente a la capacidad explicativa de la evolución. Dado que para muchos la moralidad es la esencia del ser humano, Huxley en realidad estaba diciendo que lo que nos hace humanos no podía ser abarcado por la teoría evolutiva. Sólo podemos devenir morales oponiéndonos a nuestra propia naturaleza. Fue una batida en retirada inexplicable en alguien que se había granjeado la fama de ser el «el bulldog de Darwin» por su implacable defensa de la evolución. En segundo lugar, Huxley no daba la menor pista sobre de dónde podría haber sacado la humanidad la voluntad y la fuerza para derrotar a las fuerzas de su propia naturaleza. Si en realidad somos competidores natos a los que no les preocupan los sentimientos de los demás, ¿cómo es que decidimos transformarnos en ciudadanos ejemplares? ¿Pueden las personas mantener un comportamiento atípico a lo largo de varias generaciones, como si de repente un banco de pirañas decidiera volverse vegetariano? ¿Cuán profundo puede ser un cambio de este tipo? ¿No nos convertiría esto en lobos con piel de cordero: amables por fuera y malvados por dentro?
Ésta fue la única vez en que Huxley rompió con Darwin. Como señala el biógrafo de Huxley, Adrián Desmond (1994, pág. 599): «Huxley estaba forzando su Arca ética contra la corriente darwinista que tan lejos le había permitido llegar». Dos décadas antes, Darwin había incluido de manera inequívoca la moralidad en la naturaleza humana en El origen del hombre (1982 [1871]). Se ha achacado el alejamiento de Huxley al sufrimiento que le causó la cruel mano de la naturaleza, que le arrebató la vida de su amada hija, así como a su necesidad de hacer que el despiadado cosmos darwinista resultara aceptable para el público. Al haber descrito la naturaleza como un ente implacablemente cruel y salvaje, sólo podía mantener esta postura si desplazaba la ética humana y la presentaba como una innovación independiente (Desmond, 1994). En resumen, Huxley se había puesto a sí mismo en aprietos.
El curioso dualismo de Huxley, que opone moralidad y naturaleza y humanidad frente a los demás animales, recibiría una inyección de respetabilidad gracias a los escritos de Sigmund Freud, que se basaban en los contrastes entre el consciente y el inconsciente, el ego y el superego, el Amor y la Muerte, etc. Como ocurría en el ejemplo del jardinero y el jardín de Huxley, Freud no sólo dividía el mundo en dos mitades simétricas, veía luchas por todas partes. Explicaba el tabú del incesto y otras restricciones morales como resultado de una violenta ruptura con la vida sexual espontánea de la horda primitiva, que culminaba en el sacrificio colectivo de un padre despótico a manos de sus hijos (Freud, 1962 [1913]). Dejaba que la civilización surgiera de la renuncia a los instintos, el control sobre las fuerzas de la naturaleza y la construcción de un superego cultural (Freud, 1961 [1930]).
El heroico combate de la humanidad contra las fuerzas que intentan hacerla fracasar sigue siendo en la actualidad un tema dominante en la biología, como ilustran las citas de seguidores de Huxley. Al declarar la ética como corte radical con la biología, Williams escribió sobre las miserias de la naturaleza, idea que culmina con la afirmación de que la moralidad humana es un mero producto accidental del proceso evolutivo: «Pienso que la moralidad es una cualidad accidental producida, en su estupidez sin límites, por un proceso biológico que normalmente se opone a la expresión de dicha cualidad» (Williams, 1988, pág. 438).
Después de haber explicado que nuestros genes saben lo que es mejor para nosotros y programan cada pequeño engranaje de la máquina de supervivencia humana, Dawkins esperó hasta la última frase de El gen egoísta para asegurarnos que, en realidad, podemos tirar todos esos genes por la ventana: «Somos los únicos habitantes de la Tierra que pueden rebelarse contra la tiranía de los replicadores egoístas» (Dawkins, 1976, pág. 215). La ruptura con la naturaleza es evidente en esta afirmación, como lo es la singularidad de nuestra especie. Más recientemente, Dawkins (1996) ha afirmado que somos «más buenos de lo que a nuestros genes egoístas les gustaría» y ha respaldado de forma explícita a Huxley: «Lo que estoy diciendo, al igual que muchas otras personas, entre ellos T. H. Huxley, es que tenemos derecho a expulsar el darwinismo de nuestra vida política y social, a decir que no queremos vivir en un mundo darwiniano» (Roes, 1997, pág. 3; también Dawkins, 2003).
Darwin debe estar retorciéndose en su tumba, porque el «mundo darwiniano» del que aquí se habla dista mucho del que él mismo imaginó (véase más abajo). Lo que falta en estas afirmaciones es alguna explicación sobre cómo podemos llegar a negar unos genes que esos mismos autores han descrito en otras ocasiones como todopoderosos. Como en el caso de las ideas de Hobbes, Huxley y Freud, se trata de un pensamiento por entero dualista: en lugar de un todo bien integrado, somos parte naturaleza, parte cultura. La moralidad humana se presenta como una fina corteza bajo la cual bullen pasiones antisociales, amorales y egoístas. Esta idea de la moralidad como una capa la resume bien el famoso comentario sarcástico de Ghiselin: «Arañe a un “altruista” y verá cómo sangra un “hipócrita”» (Ghiselin, 1974, pág. 247; figura 1).
Desde entonces, son muchos los divulgadores científicos que han popularizado la teoría de la capa, ente ellos Wright (1994), quien incluso llegó a afirmar que en los corazones y almas de las personas no existe la virtud y que nuestra especie es potencial pero no naturalmente moral.
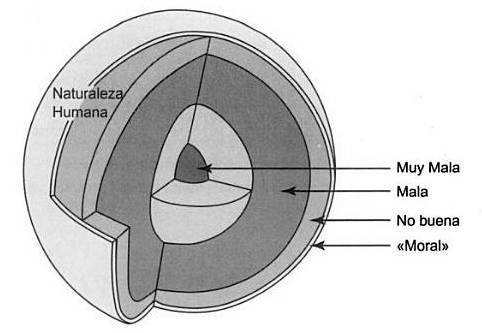
Figura 1. La visión de la moralidad sostenida por los biólogos durante el último cuarto de siglo queda resumida en la máxima de Ghiselin (1974: pág. 247): «Arañe a un “altruista” y verá cómo sangra un “hipócrita”». Se pensaba que los humanos éramos seres completamente egoístas y competitivos, y que la moralidad no era sino una ocurrencia tardía. Con el nombre de «Teoría de la capa», esta idea se remonta al contemporáneo de Darwin, Thomas Henry Huxley. Aquí se ofrece una vlsualizaclón irónica de su idea de la naturaleza humana como mala hasta su núcleo central.
Cabría entonces preguntarse: «Pero ¿qué ocurre con las personas que ocasionalmente experimentan en sí mismos y en otros cierto grado de compasión, bondad y generosidad?». Emulando a Ghiselin, Wright responde que el «animal moral» es en esencia hipócrita:
El fingimiento de egoísmo es tan común en la naturaleza humana como lo es su frecuente ausencia. Nos dotamos de un lenguaje moral elegante, negando la existencia de motivos infames y acentuando nuestra, como poco, mínima consideración por el bien superior; y criticamos duramente y en un tono de superioridad moral el egoísmo de los otros (Wright, 1994, pág. 344).
Para explicar cómo logramos vivir con nosotros mismos pese a esta farsa, los teóricos han recurrido a la noción de autoengaño. Si la gente cree que a veces es egoísta —prosigue el razonamiento— es porque están ocultándose a sí mismos sus verdaderas motivaciones (por ejemplo, Badcock, 1986). En un giro irónico de los acontecimientos, a cualquiera que no crea que nos engañamos a nosotros mismos, y que crea que la bondad verdadera existe, se le considera un pensador ilusorio y se le acusa de autoengañarse.
Algunos científicos, no obstante, han objetado:
Se dice con frecuencia que las personas respaldan estas hipótesis [sobre el altruismo humano] porque quieren que el mundo sea un lugar agradable y hospitalario. Los defensores del egoísmo y el individualismo que fomentan estas críticas practican el autohalago; se congratulan dándose palmaditas en la espalda por enfrentarse directamente con la realidad. Los egoístas e individualistas son objetivos, dicen, mientras que los defensores del altruismo y la selección grupal han caído en la trampa de una ilusión reconfortante (Sober y Wilson, 1998, págs. 8-9).
Este tira y afloja argumental sobre cómo reconciliar la bondad humana con la teoría de la evolución aparenta ser una lamentable herencia de las ideas de Huxley, quien no comprendía bien la teoría que tan eficazmente defendió frente a sus detractores. En palabras de Mayr (1997, pág. 250): «Huxley, que creía en las causas últimas, rechazaba la selección natural y en absoluto representaba el auténtico pensamiento darwinista [...]. Teniendo en cuenta lo confundido que estaba, es una pena que incluso hoy en día se le siga considerando una autoridad por su ensayo [sobre ética]».
Sin embargo, habría que señalar que en la época de Huxley ya existía una feroz oposición a sus ideas (Desmond, 1994), en parte de los biólogos rusos, como Petr Kropotkin. Dado el duro clima de Siberia, a los científicos rusos les impresionaba mucho más la lucha de los animales contra los elementos que sus luchas internas. El resultado era un énfasis en la cooperación y la solidaridad que contrastaba con la perspectiva competitiva y despiadada de Huxley (Todes, 1989). El apoyo mutuo (1972 [1902]) de Kropotkin fue un ataque contra Huxley, pero escrito con un enorme respeto por Darwin.
Aunque Kropotkin nunca formuló su teoría con la precisión y la lógica evolutiva deTrivers (1971) en su artículo seminal sobre el altruismo recíproco, ambos reflexionaron sobre los orígenes de una sociedad cooperativa, y en última instancia moral, sin invocar falsos pretextos, complejas ideas freudianas sobre la negación o forma alguna de adoctrinamiento cultural. En este sentido demostraron ser los verdaderos seguidores de Darwin.
DARWIN SOBRE LA ÉTICA
La evolución favorece a los animales que se ayudan entre sí si al hacerlo obtienen beneficios a largo plazo más valiosos que los beneficios derivados de actuar por su cuenta y competir con los demás. A diferencia de la cooperación, que se basa en beneficios simultáneos para todas las partes implicadas (conocido como mutualismo), la reciprocidad implica actos de intercambio que, aunque son beneficiosos para el receptor, son costosos para el agente (Dugatkin, 1997). Este coste, que se genera porque hay un lapso de tiempo entre dar y recibir, se elimina en cuanto se devuelve un favor de igual valor al agente (sobre el tratamiento de esta cuestión desde Trivers, 1971, véanse Axelrod y Hamilton, 1981; Rothstein y Pierotti, 1988; Taylor y McGuire, 1988). Es en estas teorías donde encontramos el germen de una explicación evolutiva de la moralidad que no tuvo en cuenta Huxley.
Es importante aclarar que estas teorías no contradicen en modo alguno las ideas al uso sobre el papel del egoísmo en la evolución. Sólo en fecha reciente se ha extraído el concepto de «egoísmo» de la lengua inglesa, despojado de su significado original, para aplicarlo fuera del terreno psicológico. Aunque para algunos el término es sinónimo de «interesado», en inglés existen diferentes términos por una razón. El egoísmo implica la intención de servirse a uno mismo, de ahí la idea de conocer lo que uno puede llegar a conseguir con un comportamiento concreto. Una planta trepadora puede desplegar un comportamiento interesado al crecer demasiado y estrangular un árbol, pero como las plantas carecen de intenciones, no pueden ser egoístas excepto en un sentido incoherente, metafórico. Por desgracia, y en una flagrante violación del significado original del término, es precisamente esta acepción vacía de significado de la palabra «egoísta» la que se ha impuesto en los debates sobre la naturaleza humana. El argumento que se escucha con frecuencia es que, si nuestros genes son egoístas, nosotros también debemos ser egoístas, pese al hecho de que los genes son simples moléculas y por tanto no pueden ser tal cosa (Midgley, 1979).
No pasa nada por describir a los animales (y a los humanos) como producto de fuerzas evolutivas que promueven el interés propio, siempre que se admita que esto en modo alguno excluye el desarrollo de tendencias altruistas y compasivas. Así lo reconoció Darwin, al explicar la evolución de estas tendencias mediante la selección grupal, en lugar de la selección individual y por parentesco que prefieren los teóricos modernos (véanse, por ejemplo, Sober y Wilson, 1998; Boehm, 1999). Darwin creía firmemente que los orígenes de la moralidad tenían perfecta cabida en sus teorías y no veía ninguna contradicción entre la dureza del proceso evolutivo y la delicadeza de algunos de sus productos. En lugar de presentar a la especie humana como un elemento exógeno a las leyes de la biología, Darwin hacía hincapié en la continuidad con los animales incluso en el terreno moral:
Cualquier animal dotado de unos instintos sociales bien marcados, incluido el cariño parental y filial, inevitablemente adquirirá un sentido moral o conciencia tan pronto como sus facultades intelectuales hayan logrado un desarrollo tan elevado, o casi tan desarrollado, como en el hombre (Darwin, 1982 [1871], págs. 71-72).
Es importante insistir en la capacidad de sentir compasión que se insinúa aquí y que Darwin expresó con más claridad en otros lugares (por ejemplo, «Muchos animales sin duda sienten compasión ante la aflicción o el peligro de otros» [Darwin, 1982 (1871), pág. 77]), porque es en este terreno donde existen sorprendentes continuidades entre los humanos y otros animales sociales. Debe de ser algo muy básico verse indirectamente afectado por las emociones de otros, porque existe constancia de estas reacciones en una gran variedad de animales y a menudo son inmediatas e incontrolables. Probablemente surgieron por primera vez con el cuidado parental, en el que se protege y alimenta a los individuos vulnerables. Sin embargo, en muchos animales estas reacciones van más allá e incluyen relaciones entre adultos no emparentados entre sí (sección 4, más abajo).
Para su idea de la compasión, Darwin se inspiró en Adam Smith, el filósofo moral escocés y padre de la economía. Dice mucho sobre las distinciones que necesitamos establecer entre el comportamiento interesado y los motivos egoístas el hecho de que Smith, famoso por su énfasis en el interés propio como principio director de la economía, escribiera también sobre el alcance universal de la compasión humana:
Por muy egoísta que pensemos que es el hombre, sin duda existen algunos principios en su naturaleza que le hacen interesarse por la fortuna de los otros y hacen que la felicidad de éstos le sea necesaria, aunque él no obtenga nada excepto el placer de verla (Smith, 1937 [1759, pág. 9).
El origen evolutivo de esta tendencia no es un misterio. Todas las especies que se sirven de la cooperación —desde los elefantes hasta los lobos y las personas— muestran lealtad al grupo y tendencias de ayuda a los demás. Estas tendencias se desarrollaron en el contexto de una vida social muy unida en la que beneficiaban a parientes y compañeros capaces de devolver un favor. Por tanto, el impulso de ayudar nunca estuvo totalmente desprovisto de un valor de supervivencia en quienes mostraban ese impulso. Pero como tantas veces ocurre, el impulso acabó por divorciarse de las consecuencias que determinaron su evolución. Esto permitió su expresión incluso cuando era improbable que se devolviera el favor, como por ejemplo cuando los beneficiarios eran desconocidos, lo que demuestra que el altruismo animal está mucho más cerca del de los humanos de lo que pensábamos y explica la llamada a que al menos temporalmente la ética deje de estar en manos de los filósofos (Wilson, 1975, pág. 562).
Personalmente, sigue sin convencerme la idea de que necesitemos la selección grupal para explicar el origen de estas tendencias; las teorías de selección por parentesco y el altruismo recíproco parecen llevarnos ya bastante lejos. Además, existe tanta migración intergrupal (de ahí el flujo genético) en los primates no humanos que no parecen darse las condiciones para la selección grupal. En todos los primates, la generación más joven de uno u otro sexo (machos en muchos monos, hembras en los chimpancés y bonobos) tiende a abandonar el grupo para unirse a grupos cercanos (Pusey y Packer, 1987). Esto significa que los grupos de primates distan mucho de estar aislados genéticamente, lo que hace poco creíble la selección grupal.
Al analizar qué constituye la moralidad, la conducta real es menos importante que las capacidades subyacentes. Por ejemplo, en lugar de sostener que compartir el alimento es un componente básico de la moralidad, son más bien las capacidades que se cree que subyacen al hecho de compartir alimento (por ejemplo, altos niveles de tolerancia, sensibilidad a las necesidad de otros, intercambio recíproco) las que resultan relevantes. También las hormigas comparten el alimento, pero probablemente sus impulsos son bastantes diferentes de los que hacen que la compartan los chimpancés o las personas (De Waal, 1989a). Darwin, que yendo más allá del comportamiento real se centró en las emociones, intenciones y capacidades subyacentes, comprendió esta diferencia. En otras palabras: la cuestión no es si los animales son o no amables entre sí, y tampoco importa mucho si su comportamiento encaja o no con nuestras preferencias morales. Lo relevante es, más bien, si poseen capacidades para la reciprocidad y la venganza, la aplicación de normas sociales, la resolución de conflictos y la compasión y la empatia (Flack y De Waal, 2000).
Esto también implica que los llamamientos a rechazar el darwinismo en nuestras vidas cotidianas para construir una sociedad moral se basan en una interpretación equivocada de Darwin. Al ver la moralidad como un producto de la evolución, Darwin imaginó un mundo mucho más habitable que el propuesto por Huxley y sus seguidores, quienes creían en una moralidad artificial y culturalmente impuesta que no recibiría ayuda alguna de la naturaleza humana. El mundo de Huxley es, con mucho, el lugar más frío y aterrador de los dos.
EDWARD WESTERMARCK
Edward Westermarck, un sueco-finés que vivió entre 1862 y 1939, merece un lugar destacado en cualquier debate sobre el origen de la moralidad, ya que fue el primer experto que promovió una visión integral que incluía tanto a los humanos como a los animales y tanto la cultura como la evolución. Es comprensible que sus ideas no fueran bien recibidas en su época, ya que iban en contra de la tradición dualista occidental que opone cuerpo y mente y cultura e instinto.
Las obras de Westermarck son una curiosa mezcla de teorías áridas, antropología pormenorizada e historias anecdóticas de animales. El autor ansiaba conectar la conducta humana y la animal, pero su propia obra se centró por completo en las personas. Dado que en aquel momento existía poca investigación sistemática sobre el comportamiento animal, tuvo que servirse de anécdotas, como la de un camello vengativo al que un camellero de 14 años había golpeado en exceso en múltiples ocasiones por rezagarse o girar por el camino equivocado. El camello aceptó el castigo pasivamente, pero, al cabo de unos días, cuando se vio sin carga y a solas en el camino con el mismo guía, «agarró la cabeza del desafortunado muchacho con su monstruosa boca, y tras levantarlo en el aire, lo volvió a arrojar al suelo con la parte superior del cráneo completamente arrancada y los sesos esparcidos por el suelo» (Westermarck, 1912 [1908], pág. 38).
No deberíamos descartar sin más estos rumores sin verificar: las historias sobre venganzas retardadas abundan en los zoológicos, sobre todo entre simios y elefantes. Ahora contamos con datos sistemáticos sobre cómo los chimpancés castigan las acciones negativas con otras acciones negativas (lo que De Waal y Luttrell, 1988, llaman un «sistema de venganza»), y sobre cómo un macaco atacado por un miembro dominante de su grupo se volvió para redirigir la agresión contra un pariente de su agresor que era más joven y vulnerable (Aureli y otros, 1992). Estas reacciones se incluyen en las emociones retributivas de Westermarck, pero para él el término «retributivas» va más allá de su connotación habitual de ajustar cuentas. También Incluye emociones positivas, como la gratitud y la devolución de servicios. Al describir las emociones retributivas como la piedra angular de la moralidad, Westermarck intervino en la cuestión del origen de la misma, anticipándose a los debates modernos sobre ética evolutiva.
Westermarck forma parte de una larga tradición que se remonta a Aristóteles y Tomás de Aquino, que ancla firmemente la moralidad en las inclinaciones y deseos naturales de nuestra especie (Arnhart, 1998, 1999). Las emociones ocupan un papel central; es bien sabido que, en lugar de ser la antítesis de la racionalidad, las emociones favorecen el razonamiento humano. Los neurocientíficos han descubierto que, por mucho que las personas razonen y reflexionen, si no hay emociones implicadas en las diferentes opciones de que disponen, nunca se alcanza una decisión o convicción (Damasio, 1994). Esto es decisivo en la elección moral, porque si hay algo que la moralidad lleve implícito, son, precisamente, las fuertes convicciones.
Estas convicciones no surgen, o más bien no pueden surgir, de la fría racionalidad, ya que requieren preocuparse por los otros y tener fuertes «instintos viscerales» sobre el bien y el mal.
Westermarck (1912 [1908], 1917 [1908]) analiza, uno por uno, toda la gama de lo que los filósofos que le precedieron, sobre todo David Hume (1985 [1739]), llamaban «sentimientos morales». Clasificó las emociones retributivas en aquellas emociones derivadas del resentimiento y la ira, que buscan la venganza y el castigo, y aquellas emociones más positivas y prosociales. Aunque en su época se conocían pocos ejemplos de emociones morales en animales —de ahí que confiara en las historias de camellos marroquíes—, ahora sabemos que existen muchos paralelismos en la conducta de los primates. También trata el concepto del «perdón» y cómo el gesto de poner la otra mejilla es apreciado universalmente. Los chimpancés se besan y abrazan después de pelearse, y estas supuestas reconciliaciones sirven para preservar la paz dentro de la comunidad (De Waal y Van Roosmalen, 1979). Existe una creciente bibliografía sobre la resolución de conflictos entre los primates y otros mamíferos (De Waal 1989b, 2000; Aureli y De Waal, 2000; Aureli y otros, 2002). La reconciliación puede no ser lo mismo que el perdón, pero sin duda ambos están relacionados.
Westermarck también ve la protección de otros frente a la agresión como el resultado de lo que él llama «resentimiento compasivo», lo que implica que este comportamiento se basa en la identificación y la empatia con el otro. La protección frente a la agresión es común en monos y simios, así como en muchos otros animales que defienden a sus parientes y amigos. La bibliografía sobre primates ofrece descripciones extensamente investigadas de coaliciones y alianzas, que algunos consideran el rasgo distintivo de la vida social de los primates y la principal razón de que los primates hayan desarrollado sociedades tan complejas y cognitivamente exigentes (por ejemplo, Byrne y Whiten, 1988; Harcourt y De Waal, 1992; De Waal, 1998 [1982]).
Del mismo modo, las emociones retributivas amables («el deseo de proporcionar placer a cambio de placer»: Westermarck, 1912 [1908], pág. 93) tienen un evidente paralelismo con lo que ahora llamamos altruismo recíproco, como la tendencia a corresponder del mismo modo a quienes nos han prestado ayuda. Westermarck añade la sanción moral como una emoción retributiva amable, de ahí que sea un componente del altruismo recíproco. Estas ideas preceden a los debates sobre la «reciprocidad indirecta» en los estudios modernos sobre ética evolutiva, que versan sobre la construcción de la reputación dentro de la comunidad (por ejemplo, Alexander, 1987). Resulta asombroso comprobar que muchas cuestiones planteadas por autores contemporáneos, expresadas en términos algo diferentes, ya están presentes en los escritos de este sueco-finés de hace un siglo.
Quizá la parte más perspicaz de la obra de Westermarck sea aquella en la que trata de abordar la cuestión de qué es lo que define a una emoción moral como moral. Aquí demuestra que en estas emociones hay algo que trasciende los puros instintos viscerales, como cuando explica que estas emociones «se diferencian de las emociones afines no morales por su desinterés, aparente imparcialidad y aire de generalidad» (Westermarck, 1917 [1908], págs. 738-739). Emociones como la gratitud y el resentimiento tienen que ver directamente con el interés propio —cómo le han tratado a uno o cómo uno desea que se le trate—, por lo que son demasiado egocéntricas para ser morales. Las emociones morales deberían estar desconectadas de la situación inmediata de uno: tratan del bien y el mal a un nivel más abstracto y desinteresado. Es sólo cuando hacemos juicios generales sobre cómo se debe tratar a alguien que podemos empezar a hablar de aprobación y desaprobación moral. Es en esta área específica, simbolizada a la perfección por el «espectador imparcial» de Smith (1937 [1759]), donde los humanos parecen llegar mucho más lejos que otros primates.
Las secciones 4 y 5 analizan la continuidad entre los dos pilares principales de la moralidad y el comportamiento de los primates. La empatía y la reciprocidad se han descrito como los principales «requisitos previos» (De Waal, 1996) o «componentes básicos» de la moralidad (Flack y De Waal, 2000) y, aunque en modo alguno son suficientes para generar la moralidad como la conocemos, son sin embargo indispensables. No cabe imaginar una sociedad moral humana sin un intercambio recíproco y un interés emocional por los otros. Esto nos brinda un punto de partida concreto para investigar la continuidad imaginada por Darwin. El debate sobre la «teoría de la capa» es fundamental en esta investigación, dado que algunos biólogos evolutivos se han desviado mucho de la idea de continuidad al presentar la moralidad como una farsa tan enrevesada que sólo existiría una especie capacitada para la misma: la nuestra. En realidad, esta opinión carece de base y, como tal, supone un obstáculo para comprender cómo devinimos morales (tabla 1). Mi intención aquí es aclarar las cosas examinando datos empíricos.
LA EMPATÍA ANIMAL
La evolución rara vez desperdicia cosas. Las estructuras se transforman, se modifican, se cooptan para otro tipo de funciones, o se «retuercen» en otra dirección: un «descenso con modificación», lo llamó Darwin. Así, las aletas frontales de los peces se transformaron en las extremidades posteriores de los animales terrestres, que a su vez se fueron transformando con el tiempo en pezuñas, garras, alas, manos y aletas. En ocasiones, una estructura determinada pierde todas sus funciones y se convierte en algo superfluo, para terminar convirtiéndose en rasgos rudimentarios sin llegar a desaparecer del todo. Así, encontramos vestigios diminutos de huesos de las piernas bajo la piel de las ballenas, o restos de pelvis en serpientes.
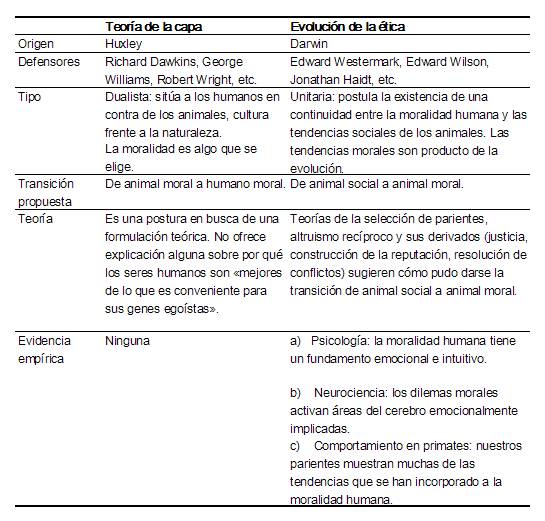
Tabla 1, Comparación entre la teoría de la capa y una visión de la moralidad como resultado de los instintos sociales.
Es por esto que para el biólogo, el modelo de muñeca rusa resulta tan satisfactorio, especialmente cuando se le dota de una dimensión histórica. Tengo una muñeca rusa que por fuera muestra al presidente Vladimir Putin, tras el cual descubrimos, por este orden, a Yeltsin, Gorbachov, Brezhnev, Kruschev, Stalin y Lenin. Para la mayoría de los analistas políticos, encontrar al pequeño Lenin o Stalin dentro de Putin no ha de ser motivo de sorpresa. Lo mismo ocurre con los rasgos biológicos: lo viejo siempre está presente en lo nuevo.
Todo esto es importante en el debate sobre el origen de la empatia, puesto que el psicólogo tiende a ver el mundo con ojos diferentes a los del biólogo. En ocasiones, los psicólogos colocan nuestros rasgos más avanzados sobre un pedestal, ignorando o incluso negando los antecedentes más sencillos de los mismos. Creen así en el cambio brusco, al menos en lo que a nuestra especie se refiere. Esto nos conduce a explicaciones poco probables sobre los orígenes que postulan discontinuidades con respecto al lenguaje, del que se dice que resulta de un «módulo» único en el cerebro humano (por ejemplo, Pinker, 1994), o con respecto a la cognición humana, de la que se dice que tiene orígenes culturales (por ejemplo, Tomasello, 1999). Es cierto que las capacidades humanas pueden alcanzar cimas verdaderamente increíbles, como por ejemplo el hecho de que yo entienda que tú me entiendes, etcétera. Pero esta «empatia reiterada», como la llaman los fenomenólogos, no es innata. Tanto desde el punto de vista del desarrollo como del de la evolución, las formas avanzadas de empatia se ven precedidas y surgen de otras formas más elementales de la misma. De hecho, bien podría decirse que las cosas son al revés. Greenspan y Shanker (2004) proponen que, más que la aparición de la lengua y la cultura en nuestra especie coincidente con el Big Bang y una posterior transformación del modo en que nos relacionamos con los demás, habría que buscar el origen de la lengua y la cultura en las tempranas conexiones emocionales y las «proto-conversaciones» que se producen entre la madre y el niño. En lugar de la empatia como meta, éste podría haber sido el punto de partida.
Los biólogos prefieren aquellas explicaciones que van de abajo arriba antes que las que van en dirección contraria, aun cuando sin duda haya espacio para estas últimas. Una vez que los procesos superiores de ordenación existen, modifican los procesos de la base. El sistema nervioso central es un buen ejemplo de este modo de procesamiento de arriba hacia abajo, tal como ocurre en el control que el córtex prefrontal ejerce sobre la memoria. La memoria no se localiza en el córtex prefrontal, pero podemos dar «órdenes» para recuperarla (Tomita y otros, 1999). Del mismo modo, la cultura y el lenguaje dan forma a las expresiones empáticas. La distinción entre «ser el origen de» y «dar forma a» es esencial, y aquí sostendré que la empatia es la forma original y prelingüística de vinculación interindividual que sólo de forma secundaria se ha visto sometida a la influencia del lenguaje y la cultura.
Las explicaciones que van de lo más simple a lo más complejo son lo opuesto de las teorías Big Bang. Presuponen una continuidad entre el pasado y el presente, entre niños y adultos, humanos y animales, incluso entre humanos y los mamíferos más primitivos. Podemos presuponer que la empatia evolucionó en primera instancia dentro del contexto del cuidado paternal, que entre los mamíferos es obligatorio (Eibl-Eibesfeldt, 1974 [1971]; MacLean, 1985). Al dar muestras de su estado mediante las sonrisas y los lloros, las crías humanas presionan a sus cuidadores para que les presten atención y actúen en consecuencia (Bowlby, 1958). Lo mismo es aplicable a otros primates. El valor de supervivencia de estas interacciones es evidente. Por ejemplo, una chimpancé hembra perdió a toda una serie de crías a pesar de su intenso y positivo interés porque estaba sorda y no corrigió los problemas en las posturas adoptadas en respuesta a sus gritos de ayuda, tales como sentarse sobre las crías o agarrarlas boca abajo (De Waal, 1998; [1982]).
En el caso de una característica humana tan omnipresente como la empatia, que además se desarrolla tan pronto (por ejemplo, Hoffman, 1975; Zahn-Waxler y Radke-Yarrow, 1990), y que muestra correlatos neurales y fisiológicos tan importantes (por ejemplo, Adolphs y otros, 1994; Rimm-Kaufman y Kagan, 1996; Decety y Chaminade, 2003), así como un sustrato genético (por ejemplo, Plomin y otros, 1993), resultaría verdaderamente extraño si no existiera una continuidad evolutiva con otros mamíferos. Sin embargo, la posibilidad de que la empatia y la compasión se den en otros animales se ha ignorado durante largo tiempo. Esto se debe en parte a un miedo excesivo al antropomorfismo, que ha sofocado los intentos de investigar las emociones animales (Pankseep, 1998; De Waal, 1999, apéndice A), y en parte también al retrato parcial que los biólogos han hecho del mundo natural como arena de combate más que de conectividad social.
¿Qué es la empatía?
Los animales sociales necesitan coordinar acciones y movimientos, responder colectivamente a situaciones de peligro, comunicarse sobre la comida y el agua, y ayudar a quienes lo necesitan. La sensibilidad o grado de respuesta a los estados de comportamiento de sus congéneres va desde la bandada de pájaros que emprende el vuelo todos a una porque uno de ellos se ha asustado ante la presencia de un predador, hasta una madre simio que vuelve hacia una cría lloriqueante para ayudarla a ir de un árbol a otro convirtiendo su cuerpo en un puente entre los dos. El primer caso es una transmisión del temor similar a un reflejo que posiblemente no implique una comprensión de lo que motivó la reacción inicial, pero que es sin lugar a dudas adaptativo. Un pájaro que nó emprenda el vuelo al mismo tiempo que el resto de la bandada podría convertirse en presa. La presión en la selección para prestar atención a los demás ha debido ser enorme. El ejemplo de la madre-simio es más selectivo, ya que implica la ansiedad de oír llorar a la propia descendencia, una evaluación de los motivos de su aflicción y un intento por mejorar la situación.
Existen numerosos ejemplos de primates que acuden en auxilio de otros en el transcurso de una pelea, rodeando con su brazo a la víctima de un ataque, u ofreciendo otras respuestas emocionales al dolor de otros (cuestión que trataremos más adelante). De hecho, se cree que prácticamente toda la comunicación entre primates no humanos está emocionalmente mediatizada. Nos resulta familiar el papel central que las emociones tienen en las expresiones faciales humanas (Ekman, 1982), pero cuando se trata de monos y simios —que cuentan con una colección de expresiones homologas (Van Hooff, 1967)— las emociones parecen igualmente importantes.
Cuando el estado emocional de un individuo hace que otro adopte un estado igual o similar, hablamos de «contagio emocional» (Hatfield y otros, 1993). Aun cuando dicho contagio es sin lugar a dudas un fenómeno básico, va más allá del hecho de que un individuo se vea afectado por el estado de otro: los dos individuos a menudo se implican en una interacción directa. Así, un niño que haya sido rechazado podrá tener una pataleta ante su madre, o un socio preferente puede mendigar comida de otro que la tenga mediante movimientos, vocalizaciones y expresiones faciales que lleven a la compasión. En otras palabras, los estados emocionales y motivaciones a menudo se manifiestan a través de comportamientos específicamente dirigidos a un compañero.
Con la creciente diferenciación entre el yo y el otro, así como una creciente apreciación de las circunstancias precisas que subyacen en los estados emocionales de los demás, el contagio emocional se convierte en empatia. La empatia comprende —y no podría haber surgido sin— el contagio emocional, pero va más allá que éste al colocar una serie de filtros entre el estado del otro y el propio. En los humanos, comenzamos a añadir estas capas cognitivas hacia los 2 años de edad aproximadamente (Eisenberg y Strayer, 1987).
Dos mecanismos relacionados con la empatia son la compasión y la angustia personal, que en sus consecuencias sociales se oponen mutuamente. La simpatía se define como «una respuesta afectiva consistente en albergar sentimientos de pesar o preocupación por otro en una situación de necesidad o angustia (más que sentir la misma emoción). Se cree que la compasión lleva implícita una motivación altruista y orientada hacia el otro» (Eisenberg, 2000, pág. 677). La angustia personal, por el contrario, hace que la parte afectada busque el alivio de su propio dolor, similar al que ha percibido en el objeto. La angustia personal no se preocupa, por tanto, de la situación de ese otro que induce a la empatia (Batson, 1990). De Waal (1996, pág. 46) ofrece un sorprendente ejemplo entre primates: los gritos de una cría de mono rhesus que haya sido duramente castigada o rechazada a menudo provoca que otras crías se aproximen, se abracen, se monten o incluso hagan una pila encima de la víctima. Así, el dolor de una cría parece extenderse a sus compañeros, que buscan posteriormente el contacto para calmar su propia excitación. En tanto que la angustia personal carece de una evaluación cognitiva y de complementareidad en la conducta, no va más allá del nivel del contagio emocional.
El hecho de que la mayoría de los libros de texto actuales sobre la cognición animal (por ejemplo, Shettleworth, 1998) no contengan en sus índices ninguna acepción dedicada a la empatia o la compasión no significa que estas capacidades no sean parte esencial de la vida de los animales; simplemente, significa que la ciencia, tradicionalmente concentrada en las capacidades individuales más que en las interindividuales, las ha pasado por alto. El empleo de herramientas y la competencia numérica, por ejemplo, son vistos como una señal de inteligencia, mientras que el trato apropiado con los demás no lo es. Es sin embargo evidente que la supervivencia a menudo depende de cómo los animales se las apañen dentro de su propio grupo, tanto en un sentido cooperativo (por ejemplo, mediante la acción concertada o la transferencia de información) como en un sentido competitivo (por ejemplo, las estrategias de dominación o el engaño). Es en el terreno de lo social, por tanto, donde uno espera encontrar los logros cognitivos más importantes. La selección debe haber favorecido aquellos mecanismos que evalúen los estados emocionales de los otros y respondan con rapidez a los mismos. La empatia es precisamente uno de esos mecanismos.
En el comportamiento humano, se da una relación muy estrecha entre empatia y compasión, y su expresión es el altruismo psicológico (por ejemplo, Hornblow, 1980; Hoffman, 1982; Batson.y otros, 1987; Eisenbergy Strayer, 1987; Wispé, 1991). Es razonable asumir que las respuestas altruistas y bondadosas de otros animales, especialmente entre los mamíferos, están basadas en mecanismos similares. Cuando Zahn-Waxler visito varios hogares con la intención de descubrir cómo los niños respondían ante miembros de su familia que habían recibido instrucciones para fingir tristeza (mediante sollozos), dolor (llorando) o angustia (fingiendo que se asfixiaban), descubrió que los niños de poco mas de 1 año ya consolaban a los demás. Dado que las expresiones de compasion emergen a una edad temprana en prácticamente todos los miembros de nuestra especie, son tan naturales como dar nuestros primeros pasos. Una consecuencia colateral de este estudio, sin embargo, fue que los animales de la casa parecían tan preocupados como los niños ante la «angustia» de los miembros de la familia. Giraban a su alrededor o ponían la cabeza en su regazo (Zahn-Waxler y otros, 1984).
Las respuestas a las emociones de los demás, enraizadas en un sentimiento de apego y en lo que Harlow denominó «el sistema afectivo» (Harlow y Harlow, 1965) se dan con frecuencia entre los animales sociales. Asi, la evidencia psicológica y de la conducta sugiere la existencia del contagio emocional en una variedad de especies (estudiadas en Preston y De Waal, 2002b, y De Waal, 2003). La interesante bibliografía escrita por psicólogos experimentales aparecida en las decadas de 1950 y 1960 colocó entre comillas términos como «empatia» y «compasión». En aquel entonces, hablar de las emociones animales era tabú. En un ensayo provocativamente titulado «Reacciones emocionales de las ratas al dolor de los otros», Church (1959) estableció que ratas que habían aprendido a apretar una palanca para conseguir comida dejaban de hacerlo si a su respuesta le acompañaba una descarga electrica que fuera visible para una rata vecina. Aun cuando esta inhibición se convirtió rápidamente en hábito, sugería cierto nivel de aversión hacia las reacciones dolorosas de los demas. Quiza tales reacciones estimularon las emociones negativas de las ratas que fueron testigos del hecho.
Los monos muestran un nivel de inhibición mayor que las ratas. La prueba más atractiva de la fuerza de la empatia en los monos la encontramos en Wechkin y otros (1964) y Masserman y otros (1964). Descubrieron que los monos rhesus se niegan a tirar de una cadena que les trae comida si con ello causan una descarga a un compañero. Un mono dejó de tirar durante cinco días y otro durante doce después de ver que uno de sus compañeros sufría una descarga. Estos monos estaban, literalmente, muriéndose de hambre con tal de evitar hacerse daño mutuamente. Un sacrificio semejante guarda relación con el estrecho sistema social y la vinculación emocional existentes entre estos macacos, como se evidencia en el hecho de que la inhibición para no dañar al otro era más pronunciada entre individuos que se conocían entre sí que entre desconocidos (Masserman y otros, 1964).
A pesar de que estos estudios tempranos sugieren que, al comportarse de determinada manera, los animales intentan aliviar o evitar el sufrimiento en los demás, no queda claro si las respuestas espontáneas hacia sus sufridos congéneres se explican mediante: a) la aversión a las señales de angustia y dolor de los otros; b) la angustia personal generada mediante contagio emocional; o c) motivaciones verdaderamente basadas en la ayuda. El trabajo con primates no humanos nos ha proporcionado más información. Algunos de los indicios son cualitativos, pero también existen datos cuantitativos sobre las reacciones de empatia.
Anécdotas para ponerse en el lugar del otro
Encontramos sorprendentes descripciones de empatia y altruismo entre primates en Yerkes (1925), Ladygina-Kohts (2000 [1935]), Goodall (1990), y De Waal (1998 [1982], 1996, 1997a). La empatia entre primates es un área tan rica que permitió a O’Connell (1995) realizar un análisis del contenido de miles de informes cualitativos. Esta investigadora llegó a la conclusión de que las respuestas al sufrimiento de otros parecen notablemente más complejas en los simios que en los monos. Para mostrar la fuerza de la respuesta empática de los simios, Ladygina-Kohts pone el ejemplo de su joven chimpancé Joni: la mejor manera de hacerle bajar del tejado de su casa (mejor que cualquier forma de castigo o recompensa) era apelando a su compasión:
Si finjo estar llorando, cierro mis ojos y sollozo; Joni inmediatamente deja de jugar o de hacer lo que esté haciendo y corre rápidamente hacia mí, muy excitado y desgreñado, desde el rincón más remoto de la casa, como por ejemplo el tejado o el techo de su jaula, de donde no podía hacerle bajar a pesar de mis persistentes ruegos para que lo hiciera. Corretea a mi alrededor con impaciencia, como si estuviera buscando al culpable; mirándome a la cara, toma con suavidad mi mentón entre sus manos, me toca la cara levemente con el dedo, como si intentara comprender qué ocurre, y se da la vuelta, apretando los dedos de los pies en forma de puño (Ladygina-Kohts, 2002 [1935], pág. 121).
De Waal (1996, 1997a) sugiere que además de la conexión emocional, los simios muestran aprecio por la situación de los demás y adoptan un cierto nivel de toma de perspectiva (apéndice B). De modo que la principal diferencia entre monos y simios no está en la empatia en sí, sino en los recubrimientos cognitivos que permiten a los simios adoptar el punto de vista del otro. En este sentido, tenemos el sorprendente ejemplo de una hembra bonobo empatizando con un pájaro en el zoo de Twycross, Inglaterra:
Un día, Kuni capturó un estornino. Temiendo que la bonobo podría molestar al aturdido pájaro, que aparentaba no haber sufrido heridas, el guardián pidió a la bonobo que lo dejara ir. Kuni cogió al estornino con una mano y escaló hasta el punto más elevado del árbol más alto, rodeando el tronco con sus piernas y así tener las dos manos libres para agarrar al pájaro. Entonces, desplegó sus alas con mucho cuidado y las abrió, un ala en cada mano, antes de arrojar al pájaro con tanta fuerza como le fue posible hacia la verja del cercado. Desgraciadamente, se quedó corta y el pájaro aterrizó a orillas del foso, donde Kuni la protegió durante largo tiempo frente a la mirada curiosa de un joven (De Waal, 1997a, pág. 156).
La acción de Kuni habría sido evidentemente inapropiada de haberla realizado con un miembro de su propia especie. Al haber visto volar a los pájaros en multitud de ocasiones, Kuni parecía haber desarrollado la noción de lo que podía ser bueno para un pájaro, ofreciendo así una versión antropoide de la capacidad para la empatia descrita de forma tan perdurable por Adam Smith (1937 [1759], pág. 10): como un «ponerse en el lugar del que sufre». Quizás el ejemplo más notable de esta capacidad sea el caso de un chimpancé que, como en los experimentos originales de la teoría de la mente de Premacky Woodruff (1978), parecía entender las intenciones de otro chimpancé y le proporcionaba asistencia específica:
Durante un invierno en el zoo de Arnhem, después de limpiar los pasillos y antes de soltar a los chimpancés, los guardianes regaron con mangueras todos los neumáticos de goma en el recinto y los fueron colgando uno a uno de un tronco horizontal que se extendía desde la estructura para la escalada. Un día, Krom se interesó por uno en el que quedaba algo de agua. Desgraciadamente, este neumático en concreto estaba al final de la hilera, con otros seis o más colgando por delante. Krom no hacía más que tirar y tirar del neumático que quería, pero no podía arrancarlo del tronco. Empujó el neumático hacia atrás, pero entonces topó con la estructura para la escalada y tampoco era posible moverlo. Krom trabajó en vano para solucionar el problema durante más de diez minutos, siendo ignorada por todos menos por Jakie, un chimpancé de 7 años de edad a quien Krom había cuidado en su infancia.
Inmediatamente después de que Krom se diese por vencida y se alejara, Jakie se aproximó. Sin dudarlo, fue sacando los neumáticos uno a uno del tronco, empezando por el que estaba delante, siguiendo por el segundo, y así sucesivamente, como haría cualquier chimpancé sensato. Cuando llegó al último neumático, lo retiró cuidadosamente para que no se perdiera ni una gota de agua, lo llevó directamente hasta su tía y lo colocó justo delante de ella. Krom aceptó su regalo sin ningún reconocimiento especial, y estaba retirando el agua con las manos cuando Jakie se marchó (adaptado de De Waal, 1996).
El hecho de que Jakie ayudara a su tía no tiene nada de raro. Lo especial es el hecho de que Jakie adivinó con exactitud lo que quería Krom. Entendió cuál era el objetivo de su tía. Esta ayuda denominada «focalizada» o de tipo selectivo es típica de los simios, pero o es rara o no se da en otros animales. Se define como un comportamiento altruista ajustado a las necesidades específicas del otro aun en situaciones novedosas, como ocurrió en el publicitado caso de Binti Jua, una gorila hembra que rescató a un niño en el zoo Brookfield de Chicago (De Waal, 1996, 1999). Un experimento reciente ha demostrado la existencia de un tipo de ayuda selectiva entre chimpancés jóvenes (Warneken y Tomasello, 2006).
Es importante señalar la importancia de la increíble fuerza de la respuesta del simio, que hace a estos animales adoptar grandes riesgos a favor de otros. Si bien en un reciente debate sobre los orígenes de la moralidad Kagan (2000) creyó obvio que un chimpancé nunca saltaría a un lago para salvar a otro, citaremos a Goodall en esta cuestión (1990, pág. 213):
En algunos zoos, los chimpancés son custodiados en islas artificiales, rodeadas de fosos llenos de agua. Los chimpancés no pueden nadar y, a menos que sean rescatados, se ahogarían si caen en aguas profundas. A pesar de esto, existen individuos que en ocasiones han realizado esfuerzos heroicos para salvar a sus compañeros, a veces con éxito. Un macho adulto perdió la vida mientras intentaba rescatar a un bebé cuya incompetente madre lo había dejado caer al agua.
Los únicos otros animales con un catálogo de respuestas similar son los delfines y los elefantes. También en este caso, las pruebas son'fundamentalmente descriptivas (para delfines, Caldwell y Caldwell, 1966; Connor y Norris, 1982; para elefantes, Moss, 1988;Payne, 1998); y aun así, resulta difícil aceptar como mera coincidencia el hecho de que los científicos que han observado a estos animales durante un período determinado de tiempo tengan un número tan elevado de ejemplos, mientras que los científicos que han observado otro tipo de animales tengan tan pocos, por no decir ninguno.
La práctica del consuelo
Esta diferencia de empatia entre monos y simios se ha visto confirmada por los estudios sistemáticos de un tipo de comportamiento conocido como «consuelo», inicialmente documentada por De Waal y Van Roosmalen (1979). Definimos la práctica del consuelo como el alivio que un espectador no involucrado ofrece a uno de los contrincantes inmersos en un incidente de agresión. Por ejemplo, un tercero acude al perdedor de una pelea y con suavidad le rodea con su brazo sobre los hombros (figura 2). El consuelo no debe confundirse con la reconciliación entre antiguos enemigos, que parece más bien motivada por el interés propio, como por ejemplo la imperiosa necesidad de restaurar una relación social alterada (De Waal, 2000). La ventaja del consuelo para el agente no es en absoluto clara. El agente podría probablemente marcharse de la escena sin consecuencias negativas.
La información sobre la práctica del consuelo entre chimpancés está bien cuantificada. De Waal y Roosmalen (1979) basaron sus conclusiones en el análisis de cientos de observaciones posconflicto, y la replicación de De Waal y Aureli (1996) incluyó un muestreo aún más amplio en el que los autores buscaban poner a prueba dos predicciones bastante sencillas. Si los contactos con terceros sirven para aliviar la angustia de los participantes en un conflicto, estos contactos deberían ir dirigidos hacia los receptores de la agresión antes que a los agresores, y en mayor medida hacia los receptores de una agresión intensa más que leve. Comparando el contacto con terceros en los niveles base, los investigadores encontraron indicios que apoyaban ambas posturas (figura 3).
Así pues, la existencia del consuelo únicamente se ha demostrado hasta el momento para el caso de los grandes simios. Cuando De Waal y Aureli (1996) se propusieron aplicar exactamente el mismo método de observación utilizado con chimpancés para detectar la práctica del consuelo en los macacos, no pudieron encontrar indicios de la misma (reseñado por Watts y otros, 2000). Esto constituyó toda una sorpresa, porque los estudios sobre la reconciliación, que básicamente utilizan el mismo método de recolección de información, han demostrado la existencia de la reconciliación en una especie tras otra. ¿Por qué, entonces, estaría la práctica del consuelo limitada a los simios?

Figura 2. Un ejemplo típico del consuelo entre chimpancés en el que un joven rodea con su brazo a un adulto que acaba de ser derrotado en una pelea con su rival. Fotografía del autor.
Es posible que la empatia cognitiva no pueda alcanzarse sin un alto grado de autoconciencia. Prestar ayuda en respuesta a situaciones concretas y a veces novedosas podría requerir una distinción entre el yo y el otro que permita que la situación del otro se divorcie de la propia, al tiempo que se mantiene el vínculo emocional que motiva el comportamiento. En otras palabras, para comprender que la fuente de la excitación vicaria no es uno mismo sino el otro, y entender las causas del estado de ese otro, es necesario establecer una clara distinción entre el otro y uno mismo.
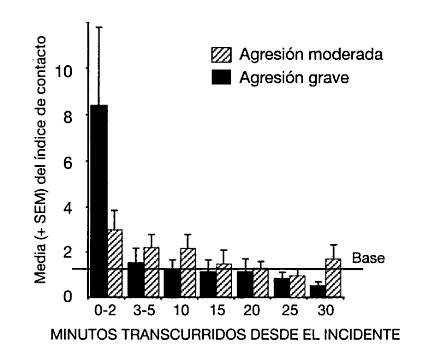
Figura 3. Frecuencia con la que terceros establecen contacto con víctimas de agresiones entre chimpancés, con una comparación entre las víctimas de agresiones graves y moderadas. Especialmente tras los primeros minutos inmediatamente posteriores al incidente, las víctimas de agresiones graves reciben más contacto que en los niveles situados en la base. Tomado de De Waal y Aureli (1996).
Basándose en estas suposiciones, Gallup (1982) fue el primero en teorizar acerca de la conexión entre la empatia cognitiva y el autorreconocimiento ante el espejo [en inglés, Mirror Self-Recognition o MSR]. Esta idea es apoyada tanto desde el punto de vista del desarrollo, debido a la correlación existente entre la emergencia del reconocimiento ante el espejo en niños pequeños y su tendencia a prestar ayuda (Bischof-Kohler, 1988; Zahn-Waxler y otros, 1992), como filogenéticamente, debido a la presencia de complejas prácticas de consuelo y ayuda entre hominoides (por ejemplo, humanos y simios), pero no entre los monos. Los hominoides son también los únicos primates capaces de autorreconocerse ante el espejo.
Anteriormente, he sostenido que además de la práctica del consuelo, la ayuda focalizada refleja la empatia cognitiva. Dicha forma de ayuda se define como un comportamiento altruista ajustado a las necesidades específicas del prójimo en situaciones nuevas, tales como la previamente descrita reacción de Kuni hacia el pájaro o el rescate de un niño por parte de Binti Jua. Estas respuestas exigen una comprensión de la situación de dificultad específica en la que se haya el individuo que precisa ayuda. Dados los indicios que apuntan a la existencia de la ayuda focalizada entre delfines (véase más arriba), el reciente descubrimiento del autorreconocimiento delante del espejo en estos mamíferos (Reiss y Marino, 2001) apoya la conexión propuesta entre una mayor autoconciencia por un lado, y la empatia cognitiva por otro.
El modelo de la muñeca rusa
La bibliografía existente incluye ejemplos de la empatia como un asunto cognitivo, hasta el punto de que los «imios, por no hablar de otros animales, probablemente carecen de ella (Povinelli, 1998; Hauser, 2000). Este punto de vista equipara la empatia a la atribución de un estado mental en los demás, y la teoría de la mente o metacognición. La postura contraria ha sido, sin embargo, defendida más recientemente en relación con los niños autistas. Frente a anteriores suposiciones de que el autismo reflejaría un déficit metacognitivo (Baron-Cohen, 2000), el autismo es perceptible mucho antes de los cuatro años, que es cuando la teoría de la mente generalmente aparece. Williams y otros (2001) sostienen que el déficit principal del autismo afecta al nivel socioafectivo, que a su vez tiene un efecto negativo sobre formas sofisticadas de percepción interpersonal, tales como la teoría de la mente o metacognición. Así, se ve la metacognición como un rasgo derivativo, cuyos antecedentes requieren una mayor atención según estos autores (postura que ahora también defiende Baron-Cohen, 2003; 2004).
Preston y De Waal (2002a) sugieren que en el centro de la capacidad para sentir empatia se encuentra un mecanismo relativamente sencillo que permite al observador (el «sujeto») acceder al estado emocional del prójimo (el «objeto») a través de las representaciones neurales y corporales del propio sujeto. Cuando el sujeto presta atención al estado del objeto, las representaciones neurales del primero de estados similares se activan automáticamente. Cuanto más cercanos y parecidos sean sujeto y objeto, más fácil será que la percepción del sujeto active respuestas motoras y autonómicas que coincidan con las del objeto (por ejemplo, cambios en el pulso cardíaco, la conductividad de la piel, la expresión facial o la postura corporal). Esta activación permite al sujeto «ponerse en la piel» del objeto, compartiendo sus sentimientos y necesidades, lo cual promueve a su vez la simpatía, la compasión y la capacidad de ayuda. El Mecanismo de Percepción-Acción (MPA) desarrollado por Preston y De Waal (2002a) concuerda con la hipótesis del marcador somático de Damasio (1994), así como con indicios más recientes sobre el vínculo en el nivel celular entre la percepción y la acción (por ejemplo, las «neuronas espejo», Di Pelligrino y otros, 1992).
La idea de que percepción y acción comparten representaciones no es nueva: se retrotrae al primer tratado sobre el Einfühlung, un concepto alemán que se tradujo al inglés como «empatia» (Wispé, 1991). Cuando Lipps (1903) hablaba de Einfühlung, que literalmente significa «sentir en», estaba especulando sobre el innere Nachahmung (o mimetismo interno) de los sentimientos ajenos en el mismo sentido propuesto por el mecanismo de percepción-acción o MPA. Así pues, la empatia es un proceso rutinario involuntario, como demuestran los estudios electromiográficos de las contracciones invisibles de los músculos faciales como respuesta a expresiones faciales humanas. Estas reacciones están plenamente automatizadas y se dan aun cuando las personas no son conscientes de lo que han visto (Dimberg y otros, 2000). Las explicaciones que ven la empatia como un proceso cognitivo superior descuidan estas reacciones instintivas, que son demasiado rápidas como para estar sometidas a un control consciente.
Los mecanismos de acción-percepción son bien conocidos en los procesos de percepción motora (Prinz and Hommel, 2002), y obliga a los investigadores a presuponer la existencia de procesos similares que subyacen en la percepción emotiva (Gállese, 2001; Wolpert y otros, 2001). Los datos sugieren que tanto la observación como la experimentación de las emociones implica una serie de sustratos psicológicos compartidos: ver el desagrado o el dolor del prójimo es muy parecido a sentirlo (Adolphs y otros, 1997, 2000; Wicker y otros, 2003). La comunicación afectiva también crea estados psicológicos parecidos en el sujeto y el objeto (Dimberg, 1982, 1990; Levenson y Reuf, 1992). En resumen, la actividad psicológica y neural humana no ocurre de forma aislada, sino que está íntimamente conectada a, y se ve afectada por, los demás seres humanos. Estudios recientes sobre la base neural de la empatia apoyan el MPA (Carr y otros, 2003; Singer y otros, 2004; De Gelder y otros, 2004).
De Waal ha descrito el modo en que las formas sencillas de la empatia se relacionan con las más complejas como una muñeca rusa (2003). Así, la empatia cubre todas las formas del estado emocional de un individuo que afectan a otros, y que contiene en su núcleo mecanismos básicos y otros mecanismos más avanzados así como habilidades cognitivas en sus capas externas (figura 4).
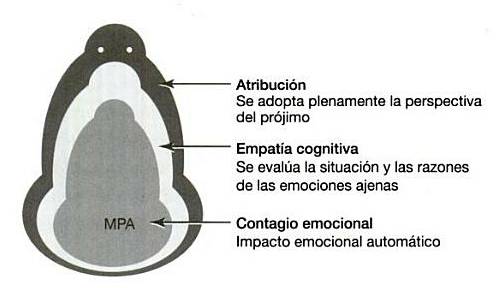
Figura 4. Según el Modelo de la Muñeca Rusa, la empatia abarca todos los procesos conducentes a los estados emocionales relacionados tanto en el sujeto como en el objeto. En su núcleo reside un Mecanismo de Percepción-Acción (MPA) que inmediatamente se traduce en una equiparación entre individuos inmediata y a menudo inconsciente de sus respectivos estados. Los niveles más elevados de la empatia que parten de esta base genéticamente programada incluyen la empatia cognitiva (por ejemplo, entender las razones de las emociones del prójimo) y la atribución del estado mental (por ejemplo, adoptar por entero la perspectiva ajena). El Modelo de la Muñeca Rusa sostiene que las capas exteriores necesitan de las Interiores. Extraído de De Waal (2003)]
El autismo podría verse reflejado en las capas externas de la muñeca rusa que estuvieran defectuosas, pero tales defectos invariablemente nos devolverían a deficiencias en las capas internas.
Esto no quiere decir que los niveles de empatia cognitivamente más elevados sean irrelevantes, pero éstos se construyen sobre esta base firme y predeterminada sin la cual estaríamos perdidos ante las motivaciones de los demás. Por supuesto, no toda la empatia puede reducirse al contagio emocional, pero no puede existir sin él. En el núcleo de esa muñeca rusa, nos encontramos con un estado emocional inducido por un mecanismo de percepción-acción (MPA) que se corresponde con el estado del objeto. En un segundo nivel, la empatia cognitiva lleva implícita una evaluación de la situación de dificultad ajena (véase De Waal, 1996). El sujeto no sólo responde a las señales que emite el objeto, sino que busca comprender las razones que le llevan a emitirlas, buscando pistas en el comportamiento y la situación del prójimo. La empatia cognitiva hace posible ofrecer un tipo de ayuda focalizada que tiene en cuenta las necesidades específicas del otro (figura 5). Estas respuestas van mucho más allá del contagio emocional, pero aun así resultarían difíciles de explicar sin la motivación proporcionada por el componente emocional. Sin él, estaríamos tan desconectados como el personaje de Mr. Spock en Star Trek, que constantemente se preguntaba por qué los demás sienten lo que dicen sentir.
Mientras que los monos (y muchos otros mamíferos sociales) parecen poseer claramente la capacidad del contagio emocional y un cierto nivel de ayuda focalizada, el segundo fenómeno no se da con tanta fuerza como entre los grandes simios. Por ejemplo, en el parque para monos Jigokudani de Japón, los guardas mantienen a las macacos primerizas alejadas de los manantiales de agua caliente porque la experiencia dice que estas hembras pueden ahogar accidentalmente a sus crías, al no prestarles atención cuando se sumergen en los estanques. Aparentemente, esto es algo que las madres mono aprenden con el tiempo; se demuestra así que no adoptan la perspectiva de su descendencia de forma automática. De Waal (1996) atribuyó su cambio de comportamiento a un «ajuste en el aprendizaje», distinguiéndolo de la empatia cognitiva que es más típica de simios y humanos. Las madres simio responden inmediata y apropiadamente a las necesidades específicas de sus crías. Por ejemplo, tienen mucho cuidado de mantenerlas alejadas del agua, y se apresuran a alejarlas de allí si se acercan.

Figura 5. La empatia cognitiva (es decir, la empatia combinada con una evaluación de la situación del prójimo) permite ofrecer un tipo de ayuda adecuada a las necesidades del otro. En este caso, una madre chimpancé extiende el brazo para ayudar a su hijo a bajar del árbol después de que éste haya gritado y se lo haya suplicado (véase la posición del brazo). Es posible que la ayuda focalizada requiera una distinción entre el yo y el otro, habilidad que también se cree que subyace en el autorreconoclmiento frente al espejo y que se encuentra en humanos, simios y delfines. Fotografía del autor.
En conclusión, la empatia no es un fenómeno que pueda ser visto en términos de blanco o negro: cubre un amplio espectro de patrones de vinculación emocional, desde los más simples y automáticos hasta los más sofisticados. Parece lógico intentar comprender en primer lugar las formas más básicas de la empatia, que de hecho están muy extendidas, antes de ocuparnos de las variaciones que la evolución cognitiva ha ido construyendo sobre esta base.
RECIPROCIDAD Y JUSTICIA
Los chimpancés y los monos capuchinos —las dos especies con las que trabajo más frecuentemente— son especiales, puesto que son de los pocos primates que comparten comida fuera del contexto madre-hijos (Feistner y McGrew, 1989). El capuchino es un primate pequeño, con el cual es fácil trabajar, a diferencia de lo que ocurre con el chimpancé, que es muchísimo más fuerte que nosotros. Los miembros de ambas especies muestran interés por la comida de la otra especie y ocasionalmente la comparten; a veces incluso se ofrecen trozos de comida entre sí. Sin embargo, la mayor parte de esta actividad compartida es pasiva, como cuando un individuo alcanza la comida que le pertenece a otro, que a su vez la deja escapar. Pero incluso el hecho de compartir de forma pasiva es especial si lo comparamos con lo que ocurre en otros animales, para quienes una situación similar tendría como resultado una pelea o una muestra de firmeza por parte del individuo dominante, sin compartir nada.
La gratitud entre los chimpancés
Estudiamos las secuencias relativas a la acción de compartir comida para ver cómo un acto beneficioso por parte de un individuo A hacia B afectaría al comportamiento de B hacia A. La tesis era que B mostraría un comportamiento beneficioso hacia A en pago por el comportamiento de éste. El problema al compartir comida, sin embargo, es que después de una sesión de prueba para todo el grupo tal como la empleamos en nuestros experimentos, la motivación para compartir cambia (los animales están saciados). De modo que el hecho de compartir no podía ser la única variable a medir. Se incluyó un segundo servicio social no afectado por el consumo de comida. Para ello, utilizamos como variable el acicalamiento entre individuos antes de compartir la comida. Medimos la frecuencia y variación de los cientos de encuentros de acicalamiento entre nuestros chimpancés por las mañanas. Transcurrida una hora y media tras estas observaciones, más o menos hacia el mediodía, dimos a los simios dos haces muy apretados de ramas y hojas. Registramos con todo detalle cerca de 7.000 interacciones con la comida, y las introdujimos en un ordenador siguiendo definiciones estrictas descritas por De Waal (1989a). La base de datos sobre servicios prestados de forma espontánea resultante excede con mucho la de cualquier otro primate no humano.
Hallamos que los adultos mostraban una mayor disposición a compartir comida con aquellos individuos que les habían acicalado con anterioridad. En otras palabras, si A había acicalado a B por la mañana, era más probable que B compartiera la comida con A más adelante. Aun así, este resultado puede tener dos explicaciones. La primera sena la hipótesis del «buen humor», según la cual aquellos individuos que han sido acicalados se encuentran en un estado de mayor benevolencia, lo que les llevaría a compartir la comida de forma indiscriminada con todos los individuos. La segunda explicación es la hipótesis del intercambio directo, según la cual el individuo que ha sido acicalado respondería compartiendo su comida directamente con el acicalador. Los datos disponibles indican que el aumento en el reparto era específico para cada acicalador. En otras palabras: los chimpancés parecían acordarse de los chimpancés que acababan de realizar un servicio (el acicalamiento) y como respuesta compartían mayores cantidades de comida con esos individuos. Asimismo, las protestas agresivas por parte de los poseedores de la comida ante los individuos que se les acercaban iban dirigidas a quienes no les habían acicalado, más que hacia quienes sí lo habían hecho. Esto constituye una prueba convincente del intercambio recíproco entre compañeros específicos (De Waal, 1997b).
De todos los ejemplos de altruismo recíproco existentes entre animales no humanos, el intercambio de comida por acicalamiento entre los chimpancés parece ser el más avanzado desde un punto de vista cognitivo. Nuestros datos apuntan con fuerza a que se trata de un mecanismo basado en la memoria. Se produjo un retraso temporal significativo entre los favores dados y los recibidos (entre media hora y dos horas); de ahí que el favor fuese correspondido mucho después de la interacción previa. Además de la memoria sobre acontecimientos pasados, debemos añadir que la memoria de un servicio recibido, como por ejemplo el acicalamiento, generó una actitud positiva hacia el individuo que había prestado el servicio, mecanismo psicológico que entre los humanos se conoce como «gratitud». YaTrivers (1971) predijo la existencia de gratitud dentro de un contexto de intercambio recíproco, idea también discutida por Bonnie y De Waal (2004). Fue clasificada por Westermarck (1912 [1908]) como una de las «emociones amables retributivas», consideradas esenciales en la moralidad humana.
El sentido de la justicia entre los monos
Es muy posible que durante la evolución de la cooperación resultase crítico que los actores comparasen sus propios esfuerzos y beneficios con los realizados y obtenidos por los demás. Las reacciones negativas podrían surgir en caso de que se violasen las expectativas. Una teoría reciente sostiene que la aversión a la desigualdad puede explicar la cooperación humana dentro de los límites del modelo de elección racional (Fehr y Schmidt, 1999). De forma parecida, las especies cooperativas no humanas parecen guiarse por una serie de expectativas relativas al resultado de la cooperación y el acceso a los recursos. De Waal (1996, pág. 95) propuso un sentido de la regularidad social, definido como «un conjunto de expectativas sobre el modo en que uno mismo (o los demás) deberían ser tratados y cómo deberían dividirse los recursos. Siempre que la realidad se desvíe en desventaja de uno mismo (o de los demás), surge una reacción negativa, comúnmente manifestada en una protesta por parte de los individuos subordinados y la práctica del castigo por parte de los individuos dominantes».
El sentido de cómo los demás deben o no deben comportarse es esencialmente egocéntrico, si bien los intereses de los individuos más próximos al actor (especialmente su familia) pueden ser tenidos en cuenta (de ahí la inclusión parentética del prójimo). Hemos de apuntar que las expectativas no han sido especificadas, sino que tienden a ser típicas de cada especie. Por ejemplo, un mono rhesüs no espera compartir la comida del individuo dominante, puesto que vive en una sociedad despóticamente jerarquizada, pero un chimpancé sí: de ahí las súplicas, los quejidos y las pataletas si no se le deja compartir. Creo que el tema de las expectativas es, de entre las cuestiones aún no estudiadas del comportamiento animal, la más importante; lo cual es aún más lamentable puesto que se trata del tema que finalmente acercará el comportamiento animal al concepto del «deber» que con tanta claridad reconocemos en el terreno de lo moral.
Al analizar las expectativas de los monos capuchinos, hicimos uso de su habilidad para juzgar y responder al valor. Sabíamos, gracias a estudios previos, que los capuchinos aprenden con facilidad a asignar un valor determinado a una muestra de agradecimiento. Es más: pueden utilizar estos valores asignados para completar un simple trueque. Esto permitió realizar un test para dilucidar la aversión a la desigualdad al medir las reacciones de los sujetos hacia un compañero que recibiera una recompensa superior al recibir los mismos objetos.
Emparejamos a cada mono con un compañero de su mismo grupo y observamos sus reacciones cuando sus compañeros recibían una recompensa mejor por realizar la misma actividad de trueque. La actividad consistía en un intercambio en el que entregábamos al sujeto un pequeño objeto que podía ser inmediatamente devuelto a cambio de una recompensa (figura 6). Cada sesión constó de veinticinco intercambios con cada individuo, y el sujeto fue testigd en todas las ocasiones del intercambio de su pareja antes del suyo. Las recompensas de comida oscilaron entre recompensas de valor inferior (como por ejemplo un trozo de pepino), por las que generalmente trabajaban con alegría, y recompensas de un valor más elevado (por ejemplo, una uva), que eran las preferidas por todos los individuos sometidos a control. Todos los sujetos fueron sometidos a: a) un Test de Equidad (TE), en el que tanto el sujeto como su compañero realizaron el mismo tipo de trabajo por el mismo tipo de comida de valor bajo; b) un Test de Desigualdad (TD), por el cual el compañero recibió una recompensa superior (la uva) por realizar el mismo esfuerzo; c) un Test de Control de Esfuerzo (TCE), diseñado para dilucidar el papel del esfuerzo, en el que el compañero recibió la uva, de más valor, gratis; y d) un Test de Control de Comida (TCC), diseñado para dilucidar el efecto de la presencia de la recompensa sobre el comportamiento del sujeto, en el que las uvas eran visibles pero no se entregaban a ningún otro capuchino.

Figura 6. Una mona capuchina en la jaula de control devuelve un pequeño objeto al responsable del experimento con su mano derecha mientras sujeta la mano humana con su mano izquierda. Su compañero la mira. Ilustración de Gwen Bragg y Frans de Waal a partir de una toma fija de vídeo.
Los individuos que recibieron recompensas de valor inferior mostraron reacciones pasivo-negativas (por ejemplo, negarse al intercambio del pequeño objeto e ignorar la recompensa) y reacciones activo-negativas. En comparación con los tests en los que ambos individuos reciben idénticas recompensas, los capuchinos se mostraban mucho menos dispuestos a completar el intercambio o aceptar la recompensa si el compañero recibía mejor trato (figura 7; Brosnan y De Waal, 2003). Los capuchinos se negaron a participar más frecuentemente si su compañero no tenía que realizar ninguna labor (un intercambio) para conseguir una recompensa mejor, que le era entregada sin esfuerzo. Evidentemente, existe siempre la posibilidad de que los sujetos estuvieran únicamente reaccionando a la presencia del alimento de mayor valor y que lo que el compañero recibía (gratis o no) no afectase a su reacción. Sin embargo, en el Test de Control de Comida, en el que la recompensa de mayor valor era visible pero no se le daba al otro mono, la reacción a la presencia de esta comida de mayor valor disminuía significativamente a lo largo del transcurso de las pruebas, lo cual supone un cambio en la dirección contraria a lo visto cuando la recompensa de mayor valor se entregaba al compañero.
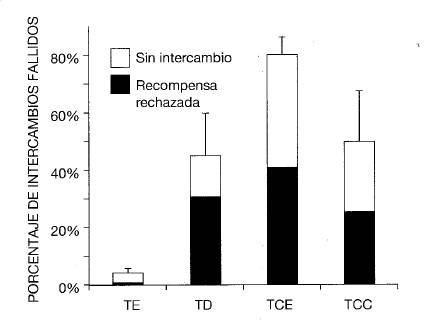
Figura 7. ± error estándar en la media del porcentaje de fracasos en el intercambio entre hembras en cuatro tipos de tests diferentes. Las barras negras representan la proporción de intercambios no realizados debido al rechazo a aceptar la recompensa; las blancas representan intercambios fallidos debido a la negativa a devolver el objeto prestado. TE: Test de Equidad, TD: Test dé Desigualdad, TCE: Test de Control de Esfuerzo, y TCC: Test de Control de Comida. El eje Y muestra el porcentaje de intercambios no realizados.
Nuestros sujetos claramente distinguían entre la comida de mayor valor consumida por un congénere y la misma comida cuando ésta era simplemente visible, intensificando su rechazo sólo en el primer caso (Brosnan y De Waal, 2003).
Los monos capuchinos parecen pues medir la recompensa en términos relativos, al comparar su recompensa con otras disponibles y sus propios esfuerzos con el de los demás. Si bien nuestros datos no permiten dilucidar las motivaciones exactas que subyacen en estas respuestas, una posibilidad es que los monos, como los humanos, se guíen por emociones sociales. En los humanos, estas emociones —conocidas como «pasiones» por los economistas— guían las reacciones individuales a la hora de realizar esfuerzos, obtener ganancias o sufrir pérdidas, y en su actitud hacia los demás (Hirschleifer, 1987; Frank, 1988; Sanfey y otros, 2003). Frente a los primates que se caracterizan por el mantenimiento de jerarquías despóticas (como los monos rhesus), es posible que las especies tolerantes con una capacidad desarrollada para la cooperación y el reparto de comida (tales como los monos capuchinos) tengan expectativas emocionales relativas a la distribución de recompensas y el intercambio social que les lleven a ver con desagrado la injusticia.
Antes de referirnos al concepto de «justicia» en este contexto conviene, no obstante, señalar una diferencia entre éste y la noción humana de justicia. Un sentido de la justicia desarrollado al máximo implicaría que una mona «rica» compartiese su comida con una «pobre», puesto que debería sentir que la compensación que recibe es excesiva. Tal comportamiento pondría de manifiesto el interés en un principio de justicia más elevado, al que Westermarck llamó (1917 [1908]) «desinteresado», y que surge de una noción verdaderamente moral de la justicia. No es éste, no obstante, el tipo de reacción que demostraron nuestros monos: su sentido de la justicia, si así podemos denominarlo, era más bien egocéntrico. Demostraron tener ciertas expectativas sobre cómo debería tratárseles, pero no sobre cómo todos los demás a su alrededor debían ser tratados. Al mismo tiempo, no puede negarse que un sentido de la justicia pleno debe tener su origen en algún punto, y que el yo es el lugar más lógico para buscar ese origen. Una vez que existe la forma egocéntrica de la justicia, puede expandirse para incluir otras formas de la misma.
MENCIO Y LA PRIMACÍA DEL AFECTO
Poco hay de nuevo bajo el sol. El énfasis puesto por Westermarck en las emociones retributivas, ya sean amistosas o de carácter vengativo, me recuerda la respuesta que Confucio ofreció a la pregunta de si existe una palabra que sirva como receta para la totalidad de la vida de una persona. Confucio propuso la palabra «reciprocidad». La reciprocidad está también, evidentemente, en el centro de la Regla de Oro, que no ha sido aún superada como el compendio de la moralidad humana. Saber que al menos parte de la psicología que subyace detrás de esta norma puede darse en otras especies junto con la empatia necesaria, refuerza la idea de que la moralidad, más que una invención reciente, es parte de la naturaleza humana.
Mencio, un seguidor de Confucio, escribió extensamente a lo largo de su vida sobre la bondad humana, entre 372 y 289 a. C. Mencio perdió a su padre a los 3 años de edad, y su madre se aseguró de que recibiera la mejor educación posible. La madre de Mencio es tan conocida como su hijo: para los chinos, sigue siendo un modelo maternal por su devoción absoluta. Conocido como el «segundo sabio» gracias a su inmensa influencia, superada solamente por Confucio, Mencio tuvo inclinaciones revolucionarias, incluso subversivas, al recalcar la obligación de los gobernantes de cubrir las necesidades del pueblo llano. Grabados en planchas de bambú y transmitidos de generación en generación a sus herederos y estudiantes, sus escritos demuestran que el debate de si somos morales por naturaleza o no viene, efectivamente, de antiguo. En un intercambio de impresiones con KaouTsze, Mencio (s.f. [372-289 a. C], págs. 270-271) reacciona frente a las ideas de este último, que nos recuerdan la metáfora del jardín y el jardinero de Huxley:
La naturaleza del hombre es como la del sauce ke; la rectitud, como una taza o un cuenco. La extracción de la benevolencia y la rectitud de la naturaleza del hombre es similar a la manufactura de tazas y cuencos a partir del sauce ke.
A lo cual Mencio replicó:
¿Es acaso posible fabricar un cuenco o una taza sin alterar la naturaleza del sauce? Debes actuar con violencia, dañar el sauce, antes de poder moldear tazas y cuencos. Si así es, entonces, según tus propios principios, ¡también sería necesario ejercer la violencia contra la humanidad y dañarla para conseguir que sea benévola y virtuosa! Tus palabras, pues, llevarían a que todos los hombres considerasen la benevolencia y la virtud una calamidad.
Mencio creía que los humanos tienden a hacer el bien de forma tan natural como el agua que corre montaña abajo. Esto queda claro en la sentencia siguiente, en la que pretende excluir la posibilidad de que exista, al más puro estilo freudiano, una doble agenda entre las motivaciones explicitadas y sentidas sobre la base de que la inmediatez de las emociones morales, tales como la compasión, no deja lugar a contorsiones cognitivas:
Cuando digo que todos los hombres poseen una mente que no les permite contemplan el sufrimiento de los demás, puede ilustrarse el significado de mis palabras de la manera que sigue: incluso hoy en día, si un grupo de hombres ve a un niño a punto de caerse en un pozo, sentirán —sin excepción— un profundo sentimiento de angustia y alarma. Y lo sentirán así no para ganarse la simpatía de los padres del niño o los elogios de amigos y vecinos, ni porque les disguste la idea de tener una reputación de seres inconmovibles ante semejante evento. De un caso como éste, podemos percibir que el sentimiento de conmiseración es esencial en el hombre (Mencio, s.f. [372-289 a. C.], pág. 78).
El ejemplo de Mencio nos recuerda al epígrafe de Westermarck («¿Podemos evitar sentir compasión por nuestros amigos?») y la cita de Smith («Por muy egoísta que supongamos al hombre...»). La idea central que subyace en las tres afirmaciones es que la angustia que sentimos al contemplar el dolor ajeno es un impulso sobre el que no ejercemos prácticamente ningún control: nos atrapa al instante, como un reflejo, sin tiempo para sopesar los pros y los contras. Las tres apuntan hacia la existencia de un proceso involuntario como mecanismo de percepción-acción (MPA). De forma notable, los posibles motivos alternativos que Mencio trae a colación figuran también en la literatura moderna, generalmente bajo el epígrafe de la construcción de la reputación. La diferencia radica, evidentemente, en que Mencio rechazó estas explicaciones por demasiado artificiales, dada la inmediatez y la fuerza del impulso compasivo. La manipulación de la opinión pública sería perfectamente posible en cualquier otro momento, afirmó, pero no en el preciso instante en el que el niño cae dentro del pozo.
Estoy absolutamente de acuerdo. La evolución ha dado lugar a especies que siguen impulsos genuinamente cooperativos. Desconozco si en el fondo la gente es buena o mala, pero creer que todas nuestras acciones están calculadas de forma egoísta —a escondidas de los demás y a menudo de nosotros mismos— equivale a sobrestimar de forma exagerada los poderes mentales del ser humano, por no hablar de los de otros animales. Más allá de los ejemplos relativos a la práctica animal del consuelo de individuos afligidos y la protección frente a las agresiones, existe una rica literatura sobre la empatia y la compasión humanas que, en líneas generales, concuerda con la estimación de Mencio de que en este ámbito los impulsos preceden a la racionalidad (por ejemplo, Batson, 1990;Wispé, 1991).
EL INTERÉS POR LA COMUNIDAD
En este ensayo, he trazado un marcado contraste entre dos escuelas de pensamiento sobre la bondad humana. Una de estas escuelas, personificada en la figura de T. H. Huxley, aún ejerce una gran influencia en nuestros días, si bien he observado que a nadie (ni siquiera entre quienes aprueban de forma explícita esta postura) le gusta que le califiquen de «teórico de la capa». Naturalmente esto puede deberse al término empleado, o al hecho de que toda vez que los supuestos que subyacen en la teoría de la capa se hacen explícitos, parece obvio que —a menos que uno esté dispuesto a seguir la vía puramente racionalista de los seguidores modernos de Hobbes, como por ejemplo Gauthier (1986)— la teoría no puede explicar cómo pasamos de ser animales amorales a ser animales morales. La teoría está reñida con la evidencia de que el procesamiento de las emociones es la fuerza que impulsa la realización de juicios morales. Si en verdad la moralidad humana pudiera reducirse a una serie de cálculos y de razonamientos, nos aproximaríamos bastante a un psicópata, que realmente no tiene ninguna intención de ser amable cuando actúa con amabilidad. La mayoría de nosotros aspira a ser algo mejor que eso, y de ahí la posible aversión a mi contraste blanquinegro entre la teoría de la capa y la corriente alternativa, que busca enraizar la moralidad en la naturaleza humana.
Esta corriente considera que la moralidad surgió de forma natural en nuestra especie, y considera que existen razones evolutivas de peso para que se desarrollasen las capacidades necesarias. Con todo, el marco teórico que explica la transición de animal social a humano moral es aún fragmentario. Encontramos sus fundamentos entre las teorías de selección de familiares y altruismo recíproco, pero resulta obvio que debemos añadir aún más elementos. Si prestamos la suficiente atención a la literatura que versa sobre la construcción de la reputación, los principios de justicia, la empatia y la resolución de conflictos (en bibliografías de índole muy diferente que no podemos reseñar aquí), parece existir un movimiento muy interesante que tiende hacia la elaboración de una teoría integrada sobre los orígenes de la moralidad (véase Katz, 2000).
Deberíamos además añadir que las presiones evolutivas responsables de nuestras tendencias morales podrían no haber sido siempre buenas o positivas. Después de todo, la moralidad es en gran medida un fenómeno intragrupal. De forma universal, los humanos tratamos a los desconocidos muchísimo peor de lo que tratamos a los miembros de nuestra propia comünidad. Es más, las normas morales apenas parecen ser aplicables fuera de nuestro entorno. Es cierto que en la época moderna existe un movimiento que busca expandir la red de la moralidad para incluir incluso a los miembros de un ejército enemigo (por ejemplo, la Convención de Ginebra, adoptada en 1949), pero todos somos conscientes de cuán frágil resulta este esfuerzo. Es muy probable que la moralidad evolucionase como un fenómeno intragrupal en conjunción con otra serie de capacidades típicamente intragrupales, tales como la resolución de conflictos, la cooperación o el acto de compartir.
No obstante, la primera forma de lealtad de los individuos no es hacia el grupo, sino hacia sí mismos y su familia. Al aumentar el nivel de interacción social y el recurso a la cooperación, los intereses compartidos debieron salir a la superficie para que la comunidad al completo se convirtiera en un aspecto importante. El paso más importante en la evolución de la moralidad humana fue la transición desde las relaciones interpersonales a un enfoque en el bien común. Entre los simios, podemos observar los comienzos de este proceso cuando solucionan conflictos ajenos. Las hembras hacen que los machos se reconcilien tras una pelea, y se convierten así en agentes de la reconciliación; los machos de mayor rango a menudo detienen las peleas entre otros individuos de forma equitativa, y así promueven la paz en el grupo. Personalmente veo este comportamiento como un reflejo de la preocupación por los intereses de la comunidad (De Waal, 1996), que a su vez refleja los intereses en juego que cada miembro del grupo tiene en el contexto de un ambiente cooperativo. La mayoría de los individuos tendría mucho que perder si la comunidad se viniera abajo, de ahí el interés por mantener la integridad y la armonía de la misma. En su estudio sobre cuestiones parecidas a ésta, Boehm (1999) añade el papel de la presión social, al menos en los humanos: toda la comunidad trabaja para recompensar el comportamiento que beneficia al grupo, y castiga aquellos comportamientos que lo socavan.
Evidentemente, la fuerza más poderosa capaz de sacar a relucir un sentido comunitario es la enemistad hacia los extraños, que obliga a que elementos que normalmente estarían enfrentados entre sí se unan. Esto puede no ser visible en el zoológico, pero es un factor muy a tener en cuenta entre los chimpancés en estado salvaje, que ejercen formas letales de violencia intercomunitaria (Wrangham y Peterson, 1996). En nuestra propia especie, nada es más evidente que nuestra tendencia a agruparnos frente a nuestros adversarios. En el transcurso de la evolución humana, la hostilidad dirigida hacia el exterior del grupo intensificó la solidaridad intragrupal, hasta el punto que hizo que surgiera la moralidad. En lugar de intentar que nuestras relaciones mejoren, como hacen los simios, hemos desarrollado enseñanzas explícitas sobre el valor de la comunidad y el lugar precedente que toma o que debe tomar sobre nuestros intereses individuales. Los humanos hemos llevado esta cuestión muchísimo más lejos que los simios (Alexander, 1987), razón por la cual nosotros tenemos sistemas morales, y ellos no.
Así pues, resulta profundamente irónico que nuestro logro más noble (la moralidad) mantenga lazos evolutivos con nuestro comportamiento más infame: la guerra. El sentimiento comunitario que la moralidad exige nos viene dado por esta última. Al traspasar el punto de encuentro entre los intereses individuales y los compartidos en conflicto, aumentamos considerablemente la presión social para asegurarnos de que todos contribuyeran al bien común.
Si aceptamos como válida esta visión de una moralidad evolucionada, es decir, de la moralidad como una consecuencia lógica de las tendencias cooperativas, al desarrollar una actitud moral y bondadosa no estaremos yendo contra nuestra naturaleza, al igual que la sociedad civil tampoco es un jardín descontrolado que tenga que ser dominado por un esforzado jardinero, como pensaba Huxley (1989 [1894]). Las actitudes morales nos han acompañado desde los comienzos de nuestra especie, y la figura del jardinero sería más bien, como muy adecuadamente la describió Dewey, la de un cultivador orgánico. Para tener éxito, el jardinero crea las condiciones adecuadas e introduce las especies vegetales que podrían no ser las normales en ese tipo de terreno «pero que entran dentro de lo que acostumbramos a encontrar en la naturaleza» (Dewey 1993 [1898], págs. 109110). En otras palabras, cuando actuamos moralmente, no engañamos de forma hipócrita a los demás: adoptamos decisiones que fluyen de unos instintos sociales más antiguos que nuestra propia especie, aun cuando les añadamos la singular complejidad humana de la preocupación desinteresada hacia los demás y hacia la sociedad en general.
A partir de la visión de Hume (1985 [1739], que consideraba a la razón esclava de las pasiones, Haidt (2001) pide una reevaluación completa del papel jugado por la racionalidad en los juicios morales, con el argumento de que la mayor parte de los actos de justificación en los humanos se dan post hoc, es decir, después de que se haya llegado a una serie de juicios morales sobre la base de intuiciones rápidas y automatizadas. Mientras que la teoría de la capa, con su énfasis en la singularidad humana, predice que la resolución de un problema moral se asigna a añadidos de nuestro cerebro evolutivamente recientes, tales como el córtex prefrontal, la neuroimagen muestra que la tarea de realizar un juicio moral implica a una gran variedad de zonas cerebrales, algunas de ellas muy antiguas (Greene y Haidt, 2002). En resumen, la neurociencia parece apoyar la postura de que la moralidad humana está evolutivamente anclada en la socialidad de los mamíferos.
Celebramos la racionalidad, pero a la hora de la verdad le asignamos un peso muy pequeño (Macintyre, 1999). Esto es especialmente cierto en el terreno de lo moral. Imaginemos que un consejero extraterrestre nos diera la orden de matar a la gente tan pronto como enfermaran de gripe. Con ello, nos diría, mataríamos un número menor de personas que el número de gente que moriría si permitiésemos que la enfermedad siguiera su curso. Al atajar la epidemia, salvaríamos vidas. Por muy lógico que esto pueda sonar, dudo que muchos optasen por este plan, debido a que la moralidad humana está firmemente anclada en las emociones sociales, con la empatia como centro. Las emociones son nuestra brújula. Matar a miembros de nuestra propia comunidad nos causa una gran repulsa, y nuestras decisiones morales son reflejo de estos sentimientos. Por esta razón, la gente se opone a poner en práctica soluciones morales que impliquen causar daño a otros (Greene y Haidt, 2002). Esto podría deberse a que la violencia siempre ha estado sujeta a la selección natural, mientras que las deliberaciones de carácter utilitario no lo han estado.
La postura intuicionista sobre la moralidad recibe apoyo de los estudios con niños. Los psicólogos del desarrollo solían creer que un niño aprende a hacer distinciones de carácter moral a raíz del miedo al castigo y del deseo de recibir elogios. Al igual que los teóricos de la capa, concebían la moralidad como procedente del exterior, algo que los adultos impondrían sobre el niño, pasivo y egoísta por naturaleza. Solía pensarse que los niños adoptaban los valores de los padres para construir el superego, la agencia moral del yo. Dejados a su libre albedrío, los niños no llegarían nunca a nada cercano a la moralidad. Sin embargo, ahora sabemos que ya a edades tempranas los niños entienden la diferencia entre los principios morales («No robes») y las convenciones culturales («No vayas en pijama a la escuela»). Aparentemente, son capaces de apreciar que la ruptura de ciertas normas hace daño y causa angustia a los demás, mientras que la ruptura de otras simplemente viola las expectativas sobre lo que es adecuado. Las actitudes de los niños no parecen estar exclusivamente basadas en nociones de castigo y recompensa. Aun cuando muchos manuales pediátricos todavía describen a los niños pequeños como monstruos egocéntricos, es evidente que al año de edad los niños ya son capaces de consolar a una persona afligida (Zahn-Waxler y otros, 1992), y que poco después comienzan a desarrollar una perspectiva moral a través de las interacciones con otros miembros de su misma especie (Killen y Nucci, 1995).
En lugar de «infligir daños al sauce», como en el ejemplo de Mencio, para hacer tazas y cuencos a partir de una moralidad artificial, nos basamos en un crecimiento natural en el que las emociones simples, como las que encontramos en los niños pequeños y animales sociales, se van desarrollando en sentimientos más refinados que incluyen a los demás y que reconocemos como subyacentes a la moralidad. Mi propia tesis aquí gira, evidentemente, alrededor de la continuidad existente entre los instintos sociales humanos y aquellos de nuestros parientes más próximos (monos y simios), pero presiento que estamos a las puertas de un giro paradigmático que terminará situando con firmeza la moralidad en el centro emocional de la naturaleza humana. Las ideas de Hume vuelven, y lo hacen a lo grande.
¿Por qué la biología evolutiva se apartó de esta senda en el último cuarto del siglo XX? ¿Por qué se consideraba la moralidad como antinatural, y por qué los altruistas eran descritos como hipócritas? ¿Por qué las emociones quedaron apartadas del debate? ¿O por qué, por ejemplo, se repetían los llamados a ir contra nuestra naturaleza y a desconfiar del «mundo darwiniano»? La respuesta se halla en lo que yo he llamado el error de Beetboven. Al igual que se dice que Ludwig van Beethoven produjo sus bellas e in tricadas composiciones en uno de los apartamentos más sucios y desordenados de toda Viena, tampoco existe una conexión entre el proceso de selección natural y sus resultados. El «error de Beethoven» consiste en pensar que, puesto que la selección natural es un proceso cruel y despiadado de eliminación, únicamente podría haber producido criaturas igualmente crueles e inmisericordes (De Waal, 2005).
Esa olla a presión que es la naturaleza, sin embargo, no funciona así. Llana y simplemente, favorece a aquellos organismos que sobreviven y se reproducen; la forma en que lo hagan es una cuestión abierta. Cualquier organismo que siendo más o menos agresivo, cooperativo o bondadoso que el resto realice la mejor tarea propagará sus genes.
En el proceso, no se especifica cuál es la receta para el éxito. La selección natural puede dar lugar a un increíble espectro de organismos, desde los más asocíales y competitivos a los más amables y benévolos. Puede que este mismo proceso no haya especificado nuestras normas y valores morales, pero nos ha dotado de la estructura psicológica, las tendencias y las habilidades necesarias para desarrollar una brújula que tenga en cuenta los intereses de la comunidad en su conjunto capaz de guiarnos en la toma de decisiones vitales. Aquí reside la esencia de la moralidad humana.