CAPÍTULO 4
DE CÓMO LA INTENCIONALIDAD SALTÓ A PRIMER PLANO
La torre de la generación y la prueba[4]
Para poder ver con anticipación en el tiempo, es útil ver más allá en el espacio. Los que fueron en un principio sistemas de monitorización internos y periféricos evolucionaron lentamente hacia sistemas capaces de discriminación no sólo proximal (cercana) sino distal (distante). Ahí es donde la percepción está a sus anchas. El sentido del olor, u olfato, se apoya en que flotan en el aire llaves precursoras para ciertas cerraduras del individuo. Las trayectorias de estos precursores son relativamente lentas, variables e inciertas a causa de la evaporación y la dispersión al azar; de este modo, se limita la información de la fuente de la cual emanan. El sentido del oído depende de que las ondas sonoras choquen con los transductores del sistema y como los caminos de las ondas del sonido son más veloces y más regulares, la percepción puede aproximarse más a una anticipación de la «acción a distancia». Pero las ondas de sonido pueden desviarse y rebotar de manera tal que puede oscurecerse su origen. La visión depende de la llegada muchísimo más rápida de fotones rebotados en las cosas del mundo, siguiendo trayectorias absolutamente rectas, de modo que con un sistema con un agujerito de forma adecuada (y, optativamente, con algunas lentes), un organismo pueda obtener información instantánea de alta fidelidad sobre los sucesos y las superficies alejados. ¿Cómo se dio esta transición de intencionalidad, de la interna a la proximal, y de la proximal a la distal? La evolución creó ejércitos de agentes internos especializados para recibir la información disponible en la periferia del cuerpo. La luz que cae sobre un pino lleva tanta información codificada como la que cae sobre una ardilla, pero ésta está provista de millones de microagentes que buscan información, específicamente diseñados para absorber, e incluso buscar, e interpretar dicha información.
Los animales no son solamente herbívoros o carnívoros. Son, según el bonito término acuñado por el psicólogo George Miller, informívoros. Y su hambre epistémica les surge de la combinación, de exquisita organización, de las hambres epistémicas específicas de millones de microagentes, organizados en docenas, o en centenares, o en miles de subsistemas. Cada uno de estos diminutos agentes puede concebirse como un sistema intencional absolutamente mínimo, cuyo proyecto de vida es hacerse una sola pregunta una y otra y otra vez: ¿Llega YA mi mensaje?, ¿llega YA mi mensaje?, y ponerse en acción de forma limitada pero adecuada todas las veces que la respuesta sea SÍ. Sin esa hambre epistémica no hay percepción, ni captación. Los filósofos han intentado a menudo analizar la percepción distinguiendo lo Dado y lo que luego hace la mente con lo Dado. Lo Dado es, por supuesto, Captado, pero esa captación de lo Dado no es cosa que haga un Gran Maestro Captador localizado en unos cuarteles generales del cerebro del animal. La tarea de captar está distribuida entre todos los captadores individualmente organizados. Los captadores no son sólo los transductores periféricos (los bastones y los conos de la retina del ojo, las células especializadas del epitelio de la nariz) sino también todos los funcionarios alimentados por éstos, células y grupos de células conectados en redes por todo el cerebro. Éstos no se alimentan de pautas luminosas o de presión (la presión de las ondas sonoras y del tacto) sino de pautas de impulsos neuronales; pero aparte de este cambio de dieta, representan papeles similares. ¿Cómo llegan a organizarse todos estos agentes en sistemas mayores capaces de sustentar formas de intencionalidad cada vez más complejas? Mediante un proceso de evolución por selección natural, desde luego, pero no mediante un proceso único.
Quiero proponer un marco en que podamos colocar las diversas opciones de diseño para el cerebro en el que veamos de dónde les viene su poder. Se trata de una estructura ultrasimplificada pero la idealización es el precio que solemos estar dispuestos a pagar para obtener una visión sinóptica. Llamo a ese marco torre de la generación y la prueba. Cada piso de la torre que se construye otorga mayor poder a los organismos de ese nivel para poder encontrar movimientos cada vez mejores y encontrarlos además con mayor eficiencia.
El poder cada vez mayor de los organismos para producir futuro puede representarse, de este modo, en una serie de pasos. Estos pasos ciertamente casi no representan con claridad períodos transitorios definidos de la historia evolutiva (no hay duda de que tales pasos los dieron diferentes linajes de manera no uniforme y solapándose unos con otros) pero los distintos pisos de la torre de la generación y la prueba marcan los avances importantes de poder cognitivo, y una vez que veamos en esbozo algunas de las características notables de cada piso, tendrá más sentido el resto de los pasos evolutivos.
En el principio fue la evolución darwiniana de las especies por selección natural. Se generó ciegamente una variedad de organismos posibles, mediante procesos de recombinación y mutación de genes más o menos arbitrarios. Se probó a estos organismos y sólo sobrevivieron los mejores diseños. Este es el piso bajo de la torre. Llamemos a sus habitantes criaturas darwinianas.
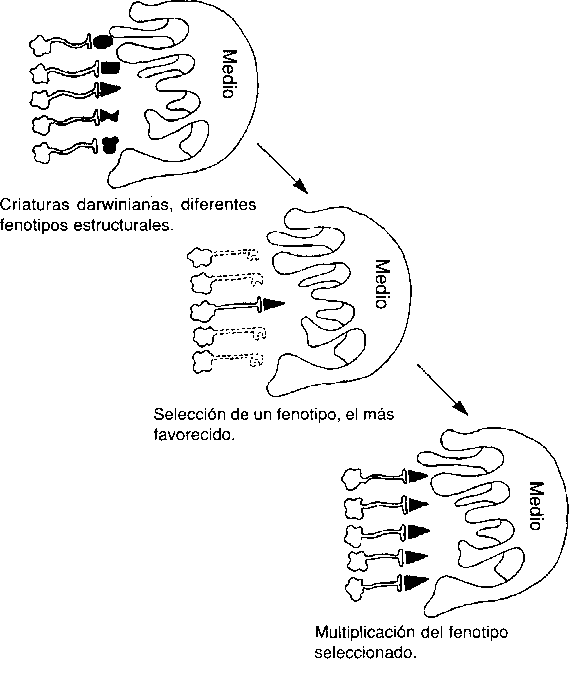
Figura 4.1
Este proceso pasó por muchos millones de ciclos, produciendo muchos diseños estupendos, tanto de plantas como de animales. En determinado momento, entre las nuevas creaciones, aparecieron algunos diseños con la propiedad de plasticidad fenotípica: es decir, que los organismos posibles individuales no estaban diseñados por completo en el momento de su nacimiento; en su diseño había elementos que podían ajustarse debido a los sucesos que se daban en las pruebas de campo. Podemos suponer que algunos de estos organismos candidatos no eran mejores que sus primos, las criaturas darwinianas acabadas, habida cuenta de que no tenían modo de favorecer (seleccionar para una repetición) las opciones de conducta para «probar las cuales» estaban equipados. Pero podemos suponer que otros tuvieron la fortuna suficiente de llevar en su interior «reforzadores» que daba la casualidad de que favorecían los movimientos inteligentes, es decir, las acciones que eran mejores para esos individuos en lugar de otras acciones disponibles. Así enfrentados al entorno, estos individuos dieron lugar a numerosas acciones, probadas una a una, hasta que encontraron la que funcionaba. Detectaron que funcionaba solamente por recibir una señal positiva o negativa del entorno, que ajustó la probabilidad de que esa acción se reprodujera en otra ocasión. Naturalmente, estarían condenadas las criaturas mal preparadas (las que tuvieran invertido el refuerzo positivo o negativo). Sólo tendrían ventaja aquellas criaturas suficientemente afortunadas por haber nacido con el refuerzo adecuado. A este subconjunto de criaturas darwinianas podemos llamarlas criaturas skinnerianas ya que como gustaba de señalar el psicólogo conductista B. F. Skinner ese «condicionante operante» no es sencillamente análogo de la selección natural darwiniana, sino una ampliación de la misma: «Allí donde cesa la conducta heredada toma el mando la modificabilidad heredada del proceso de condicionamiento».
La revolución cognitiva que surgió en la década de los setenta arrebató al conductismo su posición predominante en la psicología y desde entonces se ha dado una tendencia a subestimar el poder del condicionamiento skinneriano (o sus variantes) para conformar la competencia conductual de los organismos y dar lugar a estructuras altamente adaptativas y discernientes. Sin embargo, los florecientes trabajos de los años noventa sobre redes neurales y «conexionismo» han demostrado de nuevo el virtuosismo, a menudo sorprendente, de redes sencillas que comienzan su vida conectadas de manera más o menos al azar y que luego ajustan sus conexiones mediante una especie sencilla de «experiencia»: la historia de los refuerzos que van encontrando.

Figura 4.2
La idea fundamental de permitir al entorno interpretar un papel ciego pero selectivo en la conformación de la mente (o del cerebro, o del sistema de control) tiene un pedigrí incluso más antiguo que el de Darwin. Los antepasados intelectuales de los conexionistas de hoy y de los conductistas de ayer fueron los asociacionistas: filósofos como David Hume que en el siglo XVIII intentó imaginarse cómo las partes de la mente (partes a las que llamó impresiones e ideas) podían llegar a organizarse por sí mismas sin disponer de un director organizador que lo supiera todo. Como en una memorable ocasión me dijo un estudiante: «Hume quería que las ideas pensaran por sí mismas». Hume tuvo corazonadas maravillosas sobre cómo podrían unirse las impresiones y las ideas mediante un proceso parecido al del enlace químico creando después vías transitadas en la mente, las costumbres, pero estas corazonadas eran excesivamente vagas como para ser comprobadas. Sin embargo, el asociacionismo de Hume fue una inspiración directa para los famosos experimentos de Pavlov en el condicionamiento de la conducta animal, que a su vez llevaron a las teorías del condicionamiento, algo diferentes, de E. L. Thorndike, Skinner y demás conductistas de la psicología. Algunos de estos investigadores (concretamente, Donald Hebb) intentaron unir su conductismo todavía más a lo que se conocía del cerebro. En 1949 Hebb propuso unos modelos de mecanismos de condicionamiento simple que podrían ajustar las conexiones entre las células nerviosas. Estos mecanismos (que hoy se llaman reglas hebbianas del aprendizaje) y sus continuadores son las máquinas del cambio en el conexionismo, la última de las manifestaciones de esta tradición.
Asociacionismo, conductismo, conexionismo… en orden histórico y también en orden alfabético podemos trazar la evolución de los modelos de un tipo sencillo de aprendizaje que bien podríamos llamar aprendizaje ABC. No hay duda de que la mayoría de los animales son capaces de aprendizaje ABC, es decir, pueden llegar a modificar (o a rediseñar) su comportamiento en la dirección adecuada como resultado de un proceso de formación y de conformación debido al entorno. Hoy día existen buenos modelos, con diferente grado de realismo y de detalle, de cómo puede llegarse a semejante proceso de condicionamiento o de formación en una red de células nerviosas sin que sea una cuestión milagrosa.
Para numerosos propósitos que salvan la vida (por ejemplo, el reconocimiento de pautas, la discriminación, la generalización y el control dinámico de la locomoción) las redes ABC son francamente maravillosas: son eficientes, compactas, robustas en su funcionamiento, con un margen para los errores y relativamente fáciles de rediseñar sobre la marcha. Lo que es más, tales redes resaltan vívidamente la idea de Skinner de que hay poca diferencia entre trazar la línea entre la poda y la conformación que hace la selección natural que se transmite genéticamente a la descendencia (es decir, el cableado con el que nacemos) y la poda y la conformación que tienen lugar después en cada individuo (el recableado con el que terminamos cada uno, como resultado de la experiencia o la formación). Naturaleza y cultura se mezclan sin bordes de separación. Sin embargo, sí hay algunos trucos cognitivos que esas redes ABC no están aún preparadas para realizar y (crítica todavía más contundente) hay algunos trucos cognitivos que claramente no son resultado de formación alguna. Algunos animales parecen capaces de «aprendizaje a la primera»: pueden deducir ciertas cosas sin tener que pasar el arduo proceso de ensayo y error en el áspero mundo que es el sello de todo aprendizaje ABC.
El condicionamiento skinneriano está bien siempre que no nos mate uno de nuestros primeros errores. Un sistema mejor es el que supone una preselección entre todos los posibles comportamientos o acciones, de manera tal que los movimientos auténticamente idiotas queden fuera de las posibilidades antes de haberlos probado en la «vida real». Nosotros como seres humanos somos capaces de tal refinamiento, pero no somos los únicos. A los beneficiarios de este tercer escalón podemos denominarlos criaturas popperianas ya que, como dijo elegantemente en una ocasión el filósofo Karl Popper, esta mejora del diseño «permite que nuestras hipótesis mueran en lugar de morir nosotros». A diferencia de las criaturas meramente skinnerianas, muchas de las cuales sobreviven únicamente porque su primer movimiento es afortunado, las criaturas popperianas sobreviven porque son suficientemente listas como para hacer movimientos algo mejores que un simple movimiento afortunado. Por supuesto que tienen suerte de ser tan listas, pero ser listo es algo más que tener suerte sin más.
¿Cómo se efectúa esta preselección en los agentes popperianos? Tiene que haber un filtro y cualquier filtro de este tipo debe equivaler a una especie de entorno interno en el cual se puedan llevar a cabo con seguridad algunas pruebas: un lo que sea interno estructurado de tal forma que las acciones sustitutivas que favorece sean generalmente las mismas acciones que bendeciría también el mundo real si llegaran a realizarse. En resumidas cuentas: que el entorno interno, sea lo que sea, debe albergar montones de información del entorno externo y sus regularidades. Salvo eso (o la magia) no hay nada que pueda hacer que merezca la pena la preselección. (Siempre se puede arrojar una moneda o consultar un oráculo, pero ello no mejora la prueba a ciegas, a no ser que alguien o algo sesguen sistemáticamente tanto la moneda como el oráculo basándose en la información auténtica que del mundo poseen ese alguien o ese algo).
La belleza de la idea de Popper queda ejemplificada en el reciente desarrollo de los simuladores realistas de vuelo que se usan para entrenar a los pilotos. En un mundo simulado, los pilotos pueden aprender qué movimientos deben realizar en según qué situaciones sin arriesgar siquiera sus vidas (ni unos costosos aviones). Sin embargo, como ejemplos del truco popperiano, los simuladores de vuelo son equívocos en un aspecto: reproducen el mundo con demasiada literalidad. Debemos tener mucho cuidado de no creer que el entorno interno de una criatura popperiana es simplemente una réplica del mundo exterior, y que reproduce todas las contingencias físicas del mundo. En ese mundo milagroso de juguete ¡la estufita de nuestra cabeza tendría que estar tan caliente como para quemarnos el dedito que le ponemos encima! No hace falta suponer nada semejante. La información del efecto de colocar un dedo sobre una estufa tiene que estar ahí y debe estar de forma que pueda producir su efecto premonitorio cuando se la invoque para una prueba interna, pero tal efecto puede conseguirse sin tener que construir una réplica del mundo. Después de todo sería igual de popperiano formar a los pilotos haciéndoles leer un libro que les explicara todas las contingencias a las que podrían enfrentarse cuando se subieran a la cabina del avión. Puede que no sea un método tan poderoso de enseñanza pero ¡sería mucho mejor que salir a hacer pruebas en el aire! El elemento común de las criaturas popperianas es que de una u otra forma (bien por herencia, bien por adquisición) en ellas llevan instalada la información (información precisa del mundo que —probablemente— habrán de encontrarse) y que tal información se encuentra en forma tal que puede conseguir los objetivos que son su raison d’être.
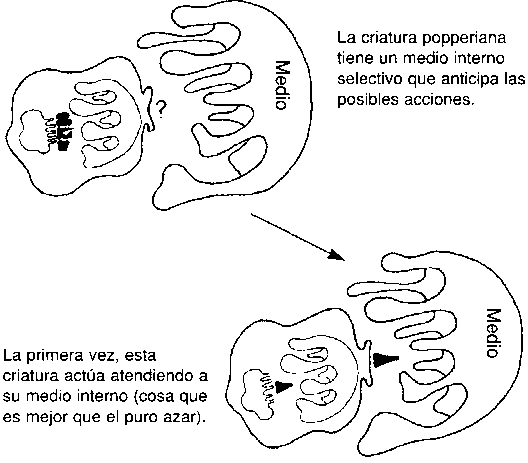
Figura 4.3
Uno de los medios por los cuales consiguen filtros útiles estas criaturas popperianas es situar las opciones posibles de conducta ante el tribunal corporal y explotar la sabiduría, por muy pasada de moda o miope que sea, acumulada en esos tejidos. Si el cuerpo se rebela (por ejemplo, con reacciones tan típicas como náusea, vértigo, temor o temblor) ya es un síntoma medianamente fiable (mejor que lanzar una moneda al aire) de que el acto que se plantea puede no ser una buena idea. Aquí se ve cómo en lugar de recablear el cerebro para eliminar tales elecciones haciéndolas estrictamente impensables, la evolución puede sencillamente organizar que la respuesta a cualquier pensamiento sobre ellas sea un agobio tan fuerte como para que sea sumamente improbable que sean las elegidas para actuar. La información del cuerpo en que se basa tal reacción puede haberse instalado allí mediante una receta genética o debido a una reciente experiencia individual. Cuando un bebé aprende a gatear tiene una aversión innata a aventurarse cerca de un panel de cristal que tenga un hueco debajo, a través del cual pueda ver un «acantilado visual». Aunque su madre le llame desde cerca, le engatuse y le anime, el niño se echa atrás temeroso pese a no haberse caído en su vida. La experiencia de sus antepasados le hace quedarse del lado seguro. Cuando una rata ha comido un nuevo tipo de alimento y se le ha inyectado una sustancia que le ha hecho vomitar, seguidamente muestra una fuerte aversión al alimento que se parece al ingerido antes de vomitar y que huele como él. Aquí la información que le induce a quedarse del lado seguro la ha obtenido de su propia experiencia. Ningún filtro es perfecto (después de todo, el panel de vidrio era seguro y el nuevo alimento de la rata no tiene nada de tóxico) pero mejor seguro que arrepentido.
Los inteligentes experimentos de psicólogos y etólogos parecen indicar algunas otras formas en las que los animales pueden probar «de cabeza» las acciones recogiendo así un beneficio popperiano. En los años treinta y cuarenta los conductistas demostraron una y otra vez que sus animales experimentales eran capaces de «aprendizaje latente» sobre el mundo, un aprendizaje que no estaba específicamente recompensado por ningún refuerzo detectable. (Este ejercicio suyo de autorrefutación es en sí un ejemplo notabilísimo de otra cuestión popperiana: que la ciencia progresa sólo cuando presenta hipótesis refutables). Cuando a las ratas se las permitía explorar un laberinto en el que no había ni comida ni ninguna otra recompensa, terminaban por aprenderse el camino en el transcurso normal de los acontecimientos; si luego se les colocaba en el laberinto algo que ellas tenían por valioso, las ratas que antes se habían aprendido el camino en incursiones anteriores lo encontraban más rápidamente (cosa nada sorprendente) que las ratas del grupo de control que veían el laberinto por primera vez. Puede parecer un descubrimiento insignificante. ¿Es que no siempre ha sido evidente que las ratas eran suficientemente listas como para aprender por dónde tenían que ir? Pues sí y no. Puede haber parecido evidente pero este es el tipo de prueba (la prueba contrastada sobre la hipótesis cero) que debe realizarse si tenemos que estar seguros de lo inteligentes que son las diversas especies y en qué medida poseen mente. Como ya veremos, otros experimentos con animales demuestran tendencias sorprendentemente estúpidas: lagunas casi increíbles en el conocimiento que los animales tienen de su propio entorno.
Valerosamente, los conductistas intentaron acomodar el aprendizaje latente en sus modelos ABC. Uno de sus recursos más contundentes era postular un «impulso de curiosidad» que se satisfacía (se «reducía» según decían ellos) mediante la exploración. Después de todo, allí había un refuerzo funcionando en todos esos medios sin refuerzo. Oh maravilla, todo entorno, por el simple hecho de ser un entorno en el que hay cosas que aprender, está lleno de estímulos de refuerzo. Como intento para salvar el conductismo ortodoxo era manifiestamente vacuo pero eso no quiere decir que sea una idea inútil en otros contextos; reconoce el hecho de que la curiosidad (hambre epistémica) debe mover cualquier sistema de aprendizaje poderoso.
Nosotros, los seres humanos, somos condicionables mediante aprendizaje ABC de modo que somos criaturas skinnerianas, pero no sólo somos criaturas skinnerianas. También disfrutamos de los beneficios de un montaje heredado genéticamente, de manera que también somos criaturas darwinianas. Pero somos más. Somos criaturas popperianas. ¿Qué otros animales son criaturas popperianas y cuáles son meramente skinnerianas? Los pichones eran los animales experimentales preferidos de Skinner, y él y sus seguidores desarrollaron la tecnología del condicionamiento operante hasta un grado sumamente sofisticado, consiguiendo que los pichones exhibieran conductas notablemente extravagantes y complejas. Cosa muy notable, los skinnerianos no han conseguido demostrar nunca que los pichones no fueran criaturas popperianas, y la investigación en un montón de especies distintas, desde pulpos hasta mamíferos, pasando por peces, parece indicar con mucha fuerza que si existen criaturas puramente skinnerianas, capaces de aprender sólo a base de aprendizaje ciego de ensayo y error, habrán de encontrarse entre los invertebrados sencillos. La gran babosa marina (o liebre marina) Aplysia californica ha reemplazado más o menos al pichón como foco de atención entre aquellos que estudian los mecanismos del condicionamiento sencillo.
Por tanto no diferimos de las demás especies en que seamos criaturas popperianas. Lejos de ello; mamíferos y pájaros, reptiles, anfibios, peces e incluso muchos invertebrados exhiben la capacidad de utilizar la información general que obtienen de sus entornos para entresacar sus opciones de conducta antes de ponerse en marcha. ¿Cómo se incorpora a sus cerebros la nueva información del entorno exterior? Evidentemente por la percepción. El entorno contiene una mezcolanza de riquezas, mucha más información de la que incluso un ángel cognitivo podría usar. Los mecanismos perceptivos diseñados para pasar por alto la mayor parte del flujo de estímulos se concentran en la información de mayor utilidad y más fiable. ¿Y cómo se las arregla la información reunida para ejercer su efecto selectivo cuando se «consideran» las opciones, ayudando al animal a diseñar interacciones todavía más efectivas con su mundo? Sin duda existe una variedad de métodos y de mecanismos diferentes, pero entre ellos se encuentran aquellos que utilizan el cuerpo como caja de resonancia.
La búsqueda de la sentiencia: informe de avances
Hemos ido añadiendo gradualmente elementos a nuestra receta de la mente. ¿Tenemos ya los ingredientes de la sentiencia? Desde luego el comportamiento normal de muchos de los animales que hemos estado describiendo aprueba nuestros intuitivos exámenes de sentiencia con un éxito rotundo. Al observar cómo tiemblan de miedo un cachorro o un bebé al borde de un aparente precipicio, o ver cómo hace muecas de aparente disgusto una rata ante el olor de un alimento supuestamente tóxico, nos vemos en dificultades incluso de mantener la hipótesis de que no estamos ante un ser sentiente. Pero también hemos dejado al descubierto algunos terrenos sólidos para la desconfianza: hemos visto algunas formas en que la conducta sorprendentemente parecida a la de una mente puede ser producida por sistemas de control relativamente sencillos, mecánicos y aparentemente ajenos a lo que sería una mente. La potencia de nuestras respuestas instintivas a la pura velocidad y al parecido vital que supone el movimiento, por ejemplo, deberían ponernos en guardia ante la posibilidad genuina, y no meramente filosófica, de vernos engañados al atribuir mayor sutileza y mayor comprensión a una entidad de lo que las circunstancias permiten. Reconociendo que el comportamiento observable puede hechizarnos, podremos apreciar la necesidad de hacernos aún más preguntas… acerca de qué hay detrás de esa conducta.
Consideremos el dolor. En 1986 el gobierno británico modificó sus leyes de protección a los animales en experimentos, añadiendo el pulpo al privilegiado círculo de animales que no pueden ser operados sin anestesia. Un pulpo es un molusco, fisiológicamente más parecido a una ostra que a una trucha (y mucho menos a un mamífero) pero la conducta del pulpo y de otros cefalópodos (calamar, sepia) es tan sorprendentemente inteligente y tan (aparentemente) sentiente que las autoridades científicas decidieron permitir que la similitud conductual superara a la diferencia interna: se supone oficialmente que los cefalópodos (pero no otros moluscos) son capaces de sentir dolor… no vaya a ser que sean capaces. Por contra, los monos rhesus están fisiológica y evolutivamente muy próximos a nosotros, de modo que solemos suponer que son capaces de sufrir como nosotros, pero llegado el momento muestran una conducta asombrosamente diferente. El primatólogo Marc Hauser me ha contado en una conversación que durante la época de apareamiento los monos machos luchan ferozmente y que no es infrecuente ver a un macho derribar a otro y luego arrancarle de un mordisco un testículo. El macho herido ni chilla ni hace muecas sino que se limita a lamerse la herida y a marcharse. Al cabo de uno o dos días ¡se le puede ver apareándose! Resulta difícil creer que este animal haya experimentado algo parecido a los dolores que experimentaría un ser humano si se le infligiera semejante herida (nos da vértigo sólo de pensarlo) a pesar de nuestra cercanía biológica. De modo que no podemos ya esperar que las pruebas fisiológicas y conductuales converjan felizmente para darnos respuestas inequívocas, habida cuenta de que ya conocemos casos en los que estos dos tipos de prueba, no concluyentes aunque irresistibles, van en sentido opuesto. ¿Cómo podemos pensar entonces en este asunto?
Una función clave del dolor es la de refuerzo negativo (el «castigo» que disminuye la probabilidad de que se repita una acción) y se puede educar a cualquier criatura skinneriana mediante refuerzo negativo de un tipo u otro. ¿Eso es lo único que es el dolor, un refuerzo negativo? ¿Dolor experimentado? ¿Podría haber dolor inconsciente o no experimentado? Hay mecanismos sencillos de refuerzo negativo que proporcionan el poder formativo o castrador de la conducta que presenta el dolor sin otros efectos secundarios de tipo mental, de modo que sería un error invocar la sentiencia siempre que nos encontremos ante un condicionamiento skinneriano. Otra función del dolor es trastornar las pautas normales de la actividad corporal que podrían empeorar una herida (por ejemplo, el dolor hace que un animal cuide su miembro herido hasta que se cure) y ello se logra mediante un flujo de sustancias neuroquímicas formando un bucle de interacciones continuadas con el sistema nervioso. Entonces ¿la presencia de esas sustancias garantiza que se da dolor? No, porque en sí mismas no son más que llaves que están flotando por ahí en busca de sus cerraduras; si se interrumpe el ciclo de interacciones no hay motivo para suponer que el dolor persista. Podemos preguntar incluso ¿son necesarias estas sustancias concretas para el dolor? ¿Podrían presentar las criaturas un sistema diferente de cerraduras y llaves? La respuesta puede depender más de los procesos históricos de evolución de este planeta que de cualquier propiedad intrínseca de las sustancias. El ejemplo del pulpo muestra que debemos mirar para ver qué variaciones pueden encontrarse en la estructura química y qué diferencias suponen en la función, pero sin esperar que estos hechos en sí mismos cierren nuestro asunto de la sentiencia.
¿Y qué pasa entonces con otros rasgos de este ciclo de interacciones? ¿En qué medida puede ser rudimentario un sistema de dolor y seguir siendo sentiencia? ¿Qué es lo que en él sería significativo y por qué? Por ejemplo, pensemos en un sapo con una pata rota. ¿Es un ser sentiente que experimenta dolor? Es un ser vivo normal cuya vida normal se ha visto interrumpida por el daño hecho a una de sus partes, impidiéndole llevar a cabo las conductas que son su modo de ganarse la vida. Todavía más: se encuentra en un estado con un potencial de refuerzo negativo muy grande: se le puede condicionar en seguida para eludir semejantes estados de su sistema nervioso. Este estado se mantiene mediante un ciclo de interacciones que en cierto modo le trastorna su normal disposición al salto… aunque en una situación de emergencia terminaría por saltar. Es tentador ver en todo esto el equivalente del dolor. Pero también es tentador atribuir al sapo un soliloquio en el cual exprese su temor de que llegue tal emergencia, ansíe un alivio, deplore su relativa vulnerabilidad, se lamente amargamente de los actos estúpidos que le han llevado a esa situación, y así sucesivamente, y todos estos añadidos no se justifican de manera alguna por lo que sabemos de los sapos. Por el contrario, cuanto más sabemos de ellos más seguros estamos de que su sistema nervioso está diseñado para que pasen por la vida sin ninguna de esas costosas capacidades reflexivas.
¿Y qué? ¿Qué tiene que ver la sentiencia con esos caprichosos talentos intelectuales? Buena pregunta, pero eso significa que debemos intentar responderla y no sencillamente utilizarla como cuestión retórica para desviar la cuestión. He aquí una circunstancia en la que nuestro modo de preguntar puede suponer una gran diferencia, porque es posible que nos engañemos creando un problema fantasma en este punto. ¿Cómo? Perdiendo la pista del lugar en el que estamos por un proceso de adición y sustracción. En el inicio, buscamos una x el ingrediente especial que distingue la mera sensibilidad de la sentiencia auténtica y trabajamos en esa búsqueda en dos direcciones. Avanzando de abajo arriba, desde los casos sencillos, añadimos versiones rudimentarias de cada rasgo por separado y en general no nos impresiona nada: aunque cada una de esas capacidades sea discutiblemente un componente esencial de la sentiencia, seguramente en la sentiencia hay más que simplemente eso: ¡un robot podría mostrar lo mismo sin tener sentiencia en absoluto! Avanzando de arriba abajo, desde nuestra propia experiencia ricamente detallada (y apreciadísima por nosotros) reconocemos que otras criaturas carecen manifiestamente de algunos de los rasgos particularmente humanos de nuestra experiencia, de manera que se los sustraemos por no ser esenciales. No queremos ser injustos con nuestros primos los animales. De tal manera que mientras nosotros admitimos que todo eso que pensamos cuando pensamos en la atrocidad del dolor (y en por qué importa moralmente que alguien sufra) supone imaginarse justamente esos añadidos antropomórficos, decidimos generosamente que son sólo añadidos no «esenciales» para el fenómeno de la sentiencia en bruto (y para su ejemplo moralmente más significativo, el dolor). Lo que podemos tender a pasar por alto, mientras estos dos barcos se cruzan en la noche, es la posibilidad de que en una vía sustraemos lo mismo que estamos buscando en la otra. Si lo que hacemos es eso, no pasaría de ser una ilusión nuestro convencimiento de que todavía tenemos que encontrar la x (el «eslabón perdido» de la sentiencia).
No digo que estemos cometiendo un error de este tipo sino que podríamos estar cometiéndolo. Es suficiente por el momento ya que altera la carga de la prueba. Por ello, he aquí una hipótesis conservadora sobre la sentiencia: no existe tal fenómeno añadido. La «sentiencia» se da en cualquier grado o intensidad imaginable desde la más sencilla y «robótica» hasta la más exquisitamente sensible e hiperreactivamente «humana». Como vimos en el capítulo 1, desde luego tenemos que trazar una línea a través de este continuo de casos multifacéticos porque es una exigencia para tener actuaciones morales, pero la perspectiva de que podamos descubrir el umbral (un «paso» que sea moralmente significativo en lo que, por lo demás, no es una escalera sino una rampa) no es que sea extremadamente improbable sino que tampoco tendrá atractivo moral.
Veamos al sapo nuevamente en este contexto. ¿De qué lado de la línea se queda el sapo? (Si los sapos son un caso excesivamente evidente para nosotros por unos u otros motivos, elijamos cualquier criatura que parezca ocupar nuestra penumbra de incertidumbre. Sea una hormiga, una medusa, una paloma o una rata). Supongamos ahora que la «ciencia confirma» que en ese sapo se encuentra la mínima sentiencia genuina: que el «dolor» del sapo es real, dolor experimentado, por ejemplo. El sapo queda ahora cualificado para el tratamiento especial reservado para los sentientes. Supongamos en vez de eso que resulta que el sapo no tiene esa x, una vez que hayamos determinado qué sea esa x. En este caso, la posición del sapo se queda en la de «mero autómata», algo en lo cual podemos intervenir de cualquier manera que se nos ocurra sin compunción moral alguna. Dado lo que ya sabemos de los sapos, ¿parece plausible que pudiera haber un rasgo desconocido hasta el momento cuyo descubrimiento pudiera justificar esa enorme diferencia en nuestra actitud? Por supuesto, si descubriéramos que los sapos son en realidad hombrecillos atrapados en cuerpos de sapo, como el príncipe del cuento, inmediatamente tendríamos la base para prodigarles los máximos cuidados porque sabríamos que, a pesar de todas sus apariencias conductuales, los sapos serían capaces de pasar por todas las torturas y las ansiedades que consideramos tan importantes en nuestro propio caso. Pero ya sabemos que el sapo no es nada semejante. Se nos ha pedido que imaginemos que existe cierta x que no es ni por asomo un príncipe humano bajo la piel de un sapo, pero que sin embargo es irresistible moralmente. No obstante, ya sabemos que un sapo no es un sencillo juego de cuerda sino una cosa viviente exquisitamente compleja capaz de una sorprendente variedad de actividades para protegerse en el fomento de su tarea prefijada de hacer más generaciones de sapos. ¿Es que esto no es suficiente para exigirnos una consideración especial por nuestra parte? Se nos pide que imaginemos que existe cierta x que no se parece en nada a esta mera complejidad de la estructura de control, pero que sin embargo nos obligaría a un reconocimiento moral cuando la descubramos. Sospecho que lo que se nos pide es que demos rienda suelta a algo más que a una fantasía. Pero prosigamos nuestra búsqueda y veamos qué viene a continuación porque aún estamos muy lejos de la mente humana.
De la fototaxia a la metafísica
Una vez que llegamos a las criaturas popperianas (criaturas cuyos cerebros tienen la capacidad de estar dotados, internamente, de habilidad preselectiva)… ¿qué viene después? Sin duda que muchas cosas diferentes, pero vamos a concentrarnos en una innovación concreta cuyos poderes podamos ver claramente. Entre los sucesores de las criaturas popperianas se encuentran aquellos cuyos entornos internos reciben la información mediante las partes diseñadas del entorno externo. Una de las ideas fundamentales más penetrantes de Darwin fue que el diseño es costoso pero que la copia de los diseños es barata; es decir, que hacer un diseño nuevo es muy difícil pero que rediseñar los diseños antiguos es relativamente sencillo. Pocos de nosotros podríamos reinventar la rueda, pero no tenemos que reinventarla ya que hemos adquirido el diseño de la rueda (y otros muchos) de la cultura en la que hemos crecido. Podemos llamar criaturas gregorianas a este sub-sub-subconjunto de criaturas darwinianas, habida cuenta de que tengo para mí que el psicólogo británico Richard Gregory es el más preeminente teórico del papel de la información (o, más exactamente, lo que Gregory llama inteligencia potencial) para la invención de movimientos inteligentes (lo que Gregory llama inteligencia cinética). Gregory observa que unas tijeras, como artefacto bien diseñado, no son sólo el resultado de una inteligencia sino de un ser dotado de inteligencia (inteligencia potencial externa) en su sentido recto y muy intuitivo: cuando le damos a alguien unas tijeras aumentamos su potencial de llegar a realizar movimientos inteligentes con mayor seguridad y más rápidamente (1981, págs. 311 y sigs.).
Los antropólogos admitieron hace tiempo que la aparición del uso de herramientas se produjo al tiempo que se daba un aumento importante de la inteligencia. Los chimpancés de la selva capturan termitas metiendo unas cañas burdamente preparadas por los termiteros, sacándolas rápidamente llenas de termitas que luego cogen pasándose la caña por la boca. Este hecho se vuelve aún más significativo cuando averiguamos que no todos los chimpancés han dado con este truco; en algunas «culturas» de los chimpancés las termitas son una fuente de alimento inexplotada. Lo cual nos recuerda que el uso de herramientas es una señal de inteligencia de ida y vuelta: no sólo requiere inteligencia reconocer y mantener una herramienta (y más aún fabricarla) sino que una herramienta confiere inteligencia a aquellos suficientemente afortunados a los que se les ha dado una. Cuanto mejor diseñada esté la herramienta (cuanta más información suponga su fabricación) mayor inteligencia potencial confiere a su usuario. Y, nos recuerda Gregory, entre las herramientas más destacadas están aquellas que él denomina herramientas mentales: las palabras.
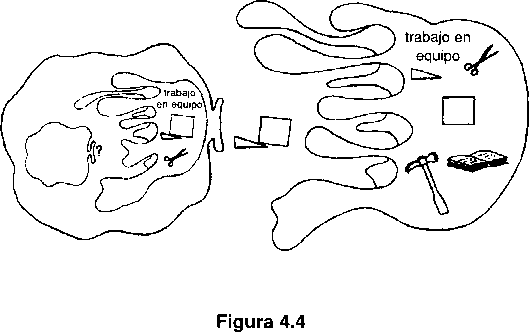
Figura 4.4
La criatura gregoriana extrae herramientas del medio (cultural): estas herramientas mejoran tanto los generadores como los probadores.
Las palabras y demás herramientas mentales proporcionan a una criatura gregoriana un entorno interior que le permite construir generadores y probadores de movimientos aún más sutiles. Las criaturas skinnerianas se preguntan: «¿Qué voy a hacer a continuación?» y no tienen clave que les dé la respuesta hasta que no se han llevado unos cuantos golpes duros. Las criaturas popperianas dan un gran paso al preguntarse: «¿Qué debo pensar a continuación?» antes de preguntarse: «¿Qué voy a hacer a continuación?». (Hay que hacer hincapié en que ni las criaturas skinnerianas ni las popperianas tienen por qué hablarse a sí mismas ni tener estos pensamientos. Sencillamente están diseñadas para funcionar como si se hubieran hecho esas preguntas. Aquí se ven el poder y el riesgo del enfoque intencional: la razón de que las criaturas popperianas sean más listas [digamos que son criaturas tortuosas con más éxito] que las skinnerianas es que responden adaptativamente a una información mayor y mejor, de un modo que podríamos describir vivida e imprecisamente a partir del enfoque intencional como si tuvieran lugar esos imaginarios soliloquios. Pero sería un error atribuir a estas criaturas todas las sutilezas que acompañan a la capacidad real de formular tales preguntas y de darse tales respuestas sobre el modelo humano de las preguntas explícitas que nos hacemos a nosotros mismos). Las criaturas gregorianas dan un gran paso hacia el nivel humano de destreza mental, beneficiándose de la experiencia de otros al explotar la sabiduría encarnada en las herramientas mentales que esos otros han inventado, mejorado y transmitido; por eso aprenden cómo pensar mejor sobre aquello que tienen que pensar a continuación… y así sucesivamente, creando una torre de sucesivas reflexiones internas sin límite fijado ni discernible. Puede verse mejor cómo llega a conseguirse este piso gregoriano volviendo la vista de nuevo a los talentos ancestrales a partir de los cuales debe de estar formada la mayoría de los talentos humanos.
Una de las prácticas más sencillas que empujan la vida y que se encuentra en muchas especies es la fototaxia: la distinción entre luz y oscuridad, orientándose hacia la primera. La luz es fácil de transducir y dado el modo en que la luz emerge de una fuente, con la intensidad disminuyendo gradualmente al alejarnos de su origen, una conexión relativamente sencilla entre transductores y efectores puede producir una fototaxia fiable. En el elegante librito del neurocientífico Valentino Braitenberg Vehicles tenemos el modelo más sencillo: el vehículo de la figura 4.5. Tiene dos transductores de luz y las señales que emiten alimentan, de forma cruzada, dos efectores (piénsese en los efectores como si fueran motores fueraborda). Cuanta más luz se transduce, más rápidamente funciona el motor. El transductor más cercano a la fuente de luz moverá su motor un poco más deprisa que el más alejado de la luz, y ello siempre haría girar el vehículo en dirección a la luz, hasta que llegara a chocar con ella u orbitara muy cerca de ella.

Figura 4.5
El mundo de un ser tan simple está graduado desde la luz hasta la oscuridad pasando por diversos grados de penumbra que recorren todo el gradiente. Este ser no sabe, ni necesita saber, nada más. El reconocimiento de la luz es casi gratuito: lo que pone en marcha al transductor es luz y al sistema le da igual si se trata de la misma luz que vuelve una y otra vez o se trata de una luz nueva. Ecológicamente, en un mundo de dos lunas sí habría diferencia en seguir una u otra: el reconocimiento de la luna sería un problema añadido que requeriría una solución. En un mundo semejante la mera fototaxia no sería suficiente. En nuestro mundo, la luna no es el tipo de objeto que una criatura tiene que saber reidentificar; con la madre suele ocurrir muchas veces justamente lo contrario.
La mamataxia (el dirigirse hacia la madre) es un talento mucho más complejo. Si mamá emitiera una luz brillante podría valer con la fototaxia pero no valdría si hubiera otras madres en las proximidades que usaran el mismo sistema. Si mamá emitiera entonces un tono concreto de luz azul, diferente de la luz que emiten todas las demás madres, entonces obtendríamos un buen resultado colocando a cada uno de nuestros fototransductores un filtro que sólo dejara pasar la luz azul. La naturaleza suele apoyarse en un principio similar pero utilizando un medio que usa la energía más eficientemente. Mamá emite un olor característico, distinguible de todos los demás olores (de las proximidades). Entonces se consigue la mamataxia (la reidentificación de la madre y la aproximación a ella) mediante la olfatotransducción u olfacción. La intensidad de los olores es función de la concentración de las llaves moleculares al difundirse en el medio circundante, el aire o el agua. Por tanto, un transductor puede ser la cerradura de forma apropiada y puede seguir el gradiente de concentración utilizando un dispositivo del tipo del vehículo de Braitenberg. Estas firmas olfativas son antiguas y poderosas. En nuestra especie se han visto sepultadas por miles de mecanismos distintos pero su situación en los cimientos sigue siendo discernible. A pesar de nuestra complejidad, los olores siguen conmoviéndonos sin que sepamos cómo o por qué, como describió en un texto ya famoso Marcel Proust[5].
La tecnología honra el mismo principio de diseño en otro medio más: el RIPE (Radiobaliza indicadora de posición de emergencia) [en inglés, EPIRB, Emergency Position Indicating Radio Beacon] un transmisor de radio autónomo y a pilas que repite una y otra vez una señal concreta en una frecuencia determinada. Se puede comprar en una tienda de efectos marineros y llevárselo uno en el barco. Si nos vemos en un apuro, lo encendemos. Inmediatamente un sistema mundial de rastreo capta nuestra señal RIPE e indica su posición con un punto luminoso en un mapa electrónico. Busca asimismo esa señal concreta en su enorme lista de señales y a partir de ella identifica nuestro barco. La identificación simplifica muchísimo la búsqueda y el rescate porque supone una redundancia: la baliza pueden rastrearla ciegamente los receptores de radio (transductores) pero conforme los rescatadores se van acercando, les ayuda saber si tienen que buscar (con la vista) un bou pesquero de color negro, un velero pequeño de color verde oscuro o una balsa de goma de color naranja brillante. Pueden añadirse otros sistemas sensoriales para hacer que la aproximación final sea más rápida y menos vulnerable a una interrupción (por ejemplo, por si se acabaran las pilas del RIPE). En los animales, el rastreo del olor no es el único medio de la mamataxia. También se depende de las señales o firmas visuales y auditivas como ha demostrado notablemente el etólogo Konrad Lorenz con sus estudios pioneros sobre las «improntas» de los gansos y patos jóvenes. Los polluelos que no reciben su impronta al poco de nacer con una firma o señal adecuada de su madre, se fijarán en la primera cosa grande que se mueva y pase por su lado y de ahí en adelante la considerarán su madre.
Las balizas (con sus complementos sensores) son buenas soluciones de diseño cuando un agente debe rastrear (reconocer, volver a identificar) a un ente concreto (normalmente otro agente, como mamá) durante largo tiempo. Se instala anticipadamente la baliza en el objetivo y adelante. (Manifestación reciente de esto son las balizas antirrobo que se instalan ocultas en los coches y se activan a distancia cuando nos han robado el coche). Pero como es normal, todo tiene su coste. Uno de los más evidentes es que amigos y enemigos pueden utilizar por igual la maquinaria de rastreo para acercarse al objetivo. Por ejemplo, es muy corriente que los predadores estén sintonizados con los mismos canales olfativos y auditivos que las crías que intentan no perder el contacto con mamá.
Olores y sonidos se emiten en cierto ámbito que no queda fácilmente bajo control del emisor. Un modo de conseguir una baliza más selectiva y de baja energía sería colocarle a mamá un punto azul concreto (un pigmento de cualquier tipo) y que la luz del sol creara en ese punto una baliza visible solamente para ciertos sectores del mundo y que desapareciera sencillamente cuando mamá se metiera en la sombra. La camada podría seguir el punto azul siempre que fuera visible. Pero este dispositivo exige una inversión en maquinaria fotosensible compleja: un ojo sencillo, por ejemplo, y no simplemente un par de fotocélulas.
La capacidad de mantenerse en estrecho y fiable contacto con alguna cosa concreta y ecológicamente muy importante (como, por ejemplo, mamá) no exige la capacidad de concebir esa cosa como una entidad concreta y duradera que va y viene. Como acabamos de ver, se puede conseguir una mamataxia fiable con unos pocos trucos. Normalmente el talento es vigoroso en los entornos sencillos pero a una criatura armada con un sistema tan simple se la «engaña» con facilidad y cuando esto ocurre se precipita hacia su desgracia sin darse cuenta de su estupidez. No hace falta que el sistema monitorice su propio éxito o reflexione sobre las condiciones bajo las cuales tiene éxito o fracasa; ese es un añadido posterior (y más costoso).
El rastreo en colaboración (en el cual el objetivo proporciona una baliza cómoda simplificando así la tarea del rastreador) es un paso en el camino hacia el rastreo competitivo, en el cual el objetivo no sólo no proporciona una baliza de firma única sino que intenta ocultarse activamente, hacerse irrastreable. Este movimiento para no ser presa se ve contrarrestado por los sistemas de rastreo general de los predadores, que lo rastrean todo, diseñados para hacer de aspectos cualesquiera que revele un objeto digno de ser rastreado una especie de baliza privada y provisional: una «imagen de búsqueda» creada para el momento por un conjunto de detectores de rasgos del predador y que se utilizan para correlacionar, a cada instante, la firma del blanco, para revisar y poner al día esa imagen de búsqueda conforme cambia el blanco y siempre con el objetivo de mantener en la retícula el objeto elegido.
Es importante admitir que esta variedad de rastreos no requiere una categorización del blanco. Pensemos en un ojo primitivo, consistente en un dispositivo de unos pocos centenares de fotocélulas que transducen un dibujo cambiante de puntos luminosos y que a su vez se activan siempre que reciben luz reflejada por algo. Un sistema así podría enviar fácilmente un mensaje de este estilo: «X, ese lo que sea que es responsable de la investigación en curso con el grupo de puntos luminosos, acaba de moverse hacia la derecha». (No es que tuviera que enviar el mensaje con tantas palabras: no harían falta en absoluto las palabras, ni símbolos, en el sistema). De modo que la única identificación que produce este sistema consiste en una especie de identificación continuada degenerada o mínima, instante tras instante, de aquello que se rastrea. Incluso aquí hay cierta tolerancia para el cambio y la sustitución. Un grupo de puntos luminosos que cambiara gradualmente y que se moviera sobre un fondo más o menos estático puede cambiar su forma y su carácter interno radicalmente y seguir siendo rastreable siempre que no cambie con demasiada brusquedad. (El fenómeno phi, en el cual el sistema visual interpreta involuntariamente las secuencias de luz destellante, como la trayectoria de un objeto en movimiento, es una manifestación vivida de este circuito que llevamos incorporado a nuestros propios sistemas de visión).
¿Qué ocurre cuando ese X se mete temporalmente detrás de un árbol? La solución más obvia es mantener intacta su imagen más reciente y luego echar un vistazo alrededor, al azar, con la esperanza de encajar otra vez esta baliza cuando aparezca, si es que aparece. Se pueden aumentar las probabilidades apuntando la imagen de búsqueda hacia el lugar más probable de reaparición de la baliza provisional. Y se puede conseguir una idea algo mejor que la puramente azarística del lugar más probable de reaparición teniendo en cuenta la trayectoria anterior de la baliza y trazando su continuación futura en línea recta. Lo cual nos proporciona ejemplos de producción de futuro en una de sus formas más antiguas y ubicuas, dándonos asimismo un caso claro de la flecha de intencionalidad apuntada hacia un blanco inexistente pero razonablemente esperable.
Esta capacidad de «mantenerse en contacto con» otro objeto (si es posible, tocándolo y manipulándolo literalmente) es el prerrequisito de la percepción de alta calidad. El reconocimiento visual de una persona o de un objeto concretos, por ejemplo, es casi imposible si la imagen del objeto no se mantiene centrada en la fóvea de alta resolución del ojo durante un tiempo apreciablemente largo. Los microagentes con hambre epistémica necesitan tiempo para alimentarse y organizarse. De tal manera que es una condición previa para desarrollar una descripción que identifique el objeto la capacidad de mantener ese foco de información acerca de una cosa concreta (de aquello, sea lo que fuere, que esté rastreando)[6].
La manera de maximizar la probabilidad de mantener o de recobrar el contacto con la entidad rastreada es confiar en múltiples sistemas independientes, todos ellos falibles pero solapándose en sus competencias. Cuando falla un sistema hay otros que lo relevan y el resultado tiende a ser un rastreo suave y continuo compuesto de elementos que funcionan intermitentemente.
¿Cómo están vinculados unos a otros estos sistemas múltiples? Hay muchas posibilidades. Si tenemos dos sistemas sensoriales, podemos unirlos mediante una compuerta Y: los dos tienen que ponerse en marcha simultáneamente con la información para que el agente responda positivamente. (Una compuerta Y puede construirse en cualquier medio: no es un objeto sino un principio organizativo. Son las dos llaves que han de abrirse para abrir una caja fuerte, o para disparar un misil nuclear: van unidas por una compuerta Y. Cuando enganchamos la manguera del jardín a un grifo por un lado y a una lanza de apertura regulable en el otro, esas dos válvulas van unidas por una compuerta Y: ambas han de estar abiertas para que salga el agua). Como alternativa, se pueden unir dos sistemas mediante una compuerta O: o uno de los dos, o A o B, o los dos al mismo tiempo, provocan una respuesta positiva del agente. Las compuertas O se usan para proporcionar un respaldo o para reservar subsistemas dentro de un sistema mayor: si falla una unidad, la actividad de la unidad de reserva es suficiente para mantener el sistema en funcionamiento. Los aviones de dos motores los llevan unidos mediante una compuerta O: lo ideal puede ser que funcionen ambos, pero en caso de necesidad basta con uno.
Conforme se van añadiendo más sistemas se perfilan maneras intermedias de unirlos. Por ejemplo, se pueden unir de manera que SI un sistema A está ACTIVADO, entonces si cualquiera de B o C está ACTIVADO, el sistema responda positivamente; en otros casos ambos sistemas, B y C, deben estar activados para producir una respuesta positiva. (Lo cual equivale a una regla de mayoría que rigiera los tres sistemas; si la mayoría, una mayoría cualquiera, está ACTIVADA, el sistema responderá positivamente). Todas las maneras posibles de unir sistemas con compuertas Y y con compuertas O (y con compuertas NO que sencillamente invierten la respuesta de un sistema, cambiando ACTIVADO por DESACTIVADO, y viceversa) se denominan funciones booleanas de esos sistemas, ya que el sistema puede describirse muy precisamente basándose en los operadores lógicos Y, O y NO, que formalizó por primera vez el matemático inglés del siglo XIX George Boole. Pero también hay maneras no booleanas de que los sistemas entremezclen sus efectos. En lugar de llevar a todos los contribuyentes a una central del voto y otorgarle a cada uno de ellos un único voto (SI o NO, ACTIVADO o DESACTIVADO) y canalizar a partir de ahí su contribución a la conducta general hasta un punto único de decisión vulnerable (el efecto sumado de todas las conexiones booleanas) podríamos permitirles que mantuvieran sus conexiones independientes y continuamente variables con la conducta y que el mundo extrajera una conducta como resultado de toda su actividad. El vehículo de Valentino Braitenberg, con sus dos fototransductores cruzados, es un ejemplo absolutamente simple de ello. La «decisión» de girar a la izquierda o a la derecha surge de la fuerza relativa de las contribuciones de los dos sistemas transductor-motor, pero el efecto no puede representarse de manera eficiente y útil como función booleana de los respectivos «argumentos» aducidos por los transductores. (En principio, el comportamiento basado en entradas y salidas de un sistema tal puede aproximarse mediante una función booleana de sus componentes si se analiza adecuadamente, pero semejante hazaña analítica puede fracasar en revelar lo que es importante en esas relaciones. Por ejemplo, considerar el tiempo atmosférico como un sistema booleano es, en principio, posible, pero eso no permite trabajar con el sistema ni obtener información de él).
Instalando docenas o centenares o miles de circuitos semejantes en un único organismo, pueden controlarse fiablemente las complejas actividades que protegen la vida, todo ello sin que ocurra nada en el interior del organismo que se parezca a tener pensamientos concretos. Se dan muchas decisiones del tipo como si: como si reconociera, como si jugara al escondite. También muchísimas maneras de que un organismo así equipado pueda «cometer errores», pero esos errores nunca equivalen a formular la representación de una proposición falsa y creer a continuación que es verdadera.
¿Qué versatilidad puede tener semejante arquitectura? Resulta difícil de decir. Los investigadores han diseñado y probado recientemente sistemas artificiales de control que producen muchas de las llamativas pautas de conducta que observamos en formas de vida relativamente sencillas, como en insectos y en otros vertebrados; de manera que es tentador creer que todas las asombrosas conductas complejas de esas criaturas puedan estar orquestadas mediante una arquitectura como esta, incluso aunque no sepamos todavía diseñar un sistema de tal complejidad. Después de todo, puede que el cerebro de un insecto no tenga más que unos cientos de neuronas y pensemos sin embargo en las complejísimas relaciones con el mundo que puede supervisar una disposición de células semejante. El biólogo evolucionista Robert Trivers indica, por ejemplo:
Las hormigas que cultivan hongos son agricultoras. Las obreras llevan hojas, las meten en los nidos, las preparan como medio de cultivo de los hongos, plantan en ellas los hongos, fertilizan los hongos con sus propias deposiciones, eliminan arrancando las especies competidoras y, por último, recolectan una parte concreta del hongo de la cual se alimentan (1985, pág. 172).
Además tenemos los rituales prologados y de pauta intrincadísima de apareo y de cría de los peces y los pájaros. Cada paso presenta exigencias sensoriales que deben satisfacerse antes de llevarse a cabo y que luego se guía adaptativamente a través de muchos obstáculos. ¿Cómo se controlan estas intrincadas maniobras? Alterando esforzadamente las fuentes de información en los experimentos, los biólogos han determinado muchas de las condiciones del entorno que se usan como indicativos aunque no es suficiente saber qué información puede captar cada organismo. La siguiente y difícil tarea es averiguar cómo pueden estar diseñados sus diminutos cerebros para dar buen uso a toda esta útil sensibilidad.
Si fuéramos un pez o un cangrejo o algo parecido y uno de nuestros proyectos consistiera en, por poner un ejemplo, construir un nido de guijarros en el fondo del océano, necesitaríamos un dispositivo para localizar guijarros y un modo de encontrar el camino de regreso a nuestro nido para depositar el guijarro hallado en un lugar adecuado antes de seguir buscando. Sin embargo, no hace falta que el sistema sea a prueba de estupideces. Como es improbable que, durante nuestras búsquedas, surjan subrepticiamente nidos de guijarros impostores en la zona que hayamos elegido (por lo menos hasta que los listos experimentadores humanos se toman interés en nosotros), podemos mantener unos niveles de reidentificación bastante bajos y poco costosos. Si se da un error en la «identificación», probablemente seguiríamos construyendo, no engañados por la treta sino absolutamente incapaces de reconocer o de apreciar el error, es decir, sin que nos preocupara lo más mínimo. Por otra parte, si estuviéramos equipados con un sistema de respaldo para el reconocimiento del nido, y el nido impostor no superara la prueba de respaldo nos veríamos metidos en el desorden, cogidos entre dos sistemas que nos requieren al mismo tiempo. Estos conflictos se dan pero no tiene sentido preguntar cuando el tal organismo va nervioso de un lado para otro: «¿Y ahora, qué está pensando? ¿Cuál es el contenido proposicional de su estado de confusión?».
En organismos como el nuestro (organismos equipados con muchísimas capas de sistemas que se controlan a sí mismos, que pueden comprobar la existencia de estos conflictos cuando se presentan y pueden intentar mediar en ellos) a veces queda perfectamente claro qué error se ha cometido. Un ejemplo perturbador es la ilusión de Capgras, una afección extravagante que a veces padecen los seres humanos que han sufrido una lesión cerebral. La señal distintiva de la ilusión de Capgras es el convencimiento del paciente de que un conocido cercano (generalmente un ser querido) se ha visto suplantado por un impostor que se le parece (y que habla y actúa como él) y que el conocido ¡ha desaparecido misteriosamente! Este sorprendente fenómeno debería agitar las aguas de la filosofía. Los filósofos han reunido muchos casos extraños de identidad cambiada para ilustrar sus diversas teorías filosóficas y la literatura de la filosofía está plagada de fantásticos experimentos mentales sobre espías y asesinos que viajan de incógnito, amigos íntimos que se disfrazan de gorila y gemelos idénticos que se reencuentran al cabo de muchísimo tiempo, pero los casos de la vida real de la ilusión de Capgras no han llamado todavía la atención de los filósofos. Lo que es particularmente llamativo de estos casos es que no dependen de sutiles disfraces ni de atisbos fugaces. Al contrario: la ilusión persiste incluso cuando el agente somete al individuo que sirve de blanco a un estudio detallado. Se sabe de pacientes de Capgras que han asesinado a sus cónyuges ¡tan seguros estaban de que aquellos intrusos estaban intentando meterse en pellejos (en vidas al completo) que no les pertenecían por derecho propio! No puede haber duda de que en semejante caso, tan triste, el agente en cuestión ha dado por ciertas determinadas proposiciones específicas de no identidad: este hombre no es mi marido; este hombre es cualitativamente lo más parecido a mi marido y sin embargo no es mi marido. Resulta muy interesante el hecho de que las personas que sufren de semejante ilusión son claramente incapaces de decir por qué están tan seguras.
El neuropsicólogo Andrew Young (1994) ofrece una hipótesis ingeniosa y plausible para explicar qué es lo que va mal. Young contrasta la ilusión de Capgras con otra curiosa afección causada por lesiones cerebrales: la prosopagnosia. Las personas con prosopagnosia no pueden reconocer rostros humanos familiares. Pueden tener buena vista pero no son capaces de identificar ni siquiera a sus amigos íntimos hasta que les oyen hablar. En un experimento característico se les muestran colecciones de fotografías: algunas son de individuos desconocidos y otras de miembros de su familia y personajes conocidos: Hitler, Marilyn Monroe, J. F. Kennedy. Cuando se les pide que cojan las de rostros familiares, no lo hacen mejor que si las escogieran al azar. Pero durante más de una década los investigadores han sospechado que a pesar de un resultado tan asombrosamente pobre, algo había en algunos prosopagnósicos que identificaba correctamente a los miembros de la familia y a la gente famosa porque su cuerpo reaccionaba de manera diferente ante las caras familiares. Si mientras miran la foto de una cara conocida se les dicen diversos nombres posibles de la persona fotografiada, muestran una alta respuesta galvánica de la piel al oír el nombre correcto. (La respuesta galvánica de la piel es la medida de la conductividad eléctrica y la principal prueba en la que se basan los polígrafos o «detectores de mentiras»). La conclusión que Young y otros investigadores sacan de estos resultados es que debe haber dos (o más) sistemas que pueden identificar un rostro y que uno de ellos está intacto en los prosopagnósicos que muestran esta respuesta. Este sistema sigue funcionando bien, furtiva y prácticamente sin ser notado. Supongamos ahora, dice Young, que los pacientes de Capgras tengan la disfuncionalidad opuesta: el sistema (o sistemas) que opera al descubierto para el reconocimiento consciente de rostros funciona perfectamente (y por eso los pacientes de Capgras están de acuerdo en que los «impostores» tienen el mismo aspecto que sus seres queridos) pero que el sistema (o sistemas) furtivo que normalmente proporciona un voto de confianza al reafirmar lo visto en esas ocasiones está deteriorado y ominosamente silencioso. La ausencia de tal colaboración sutil a la identificación es tan irritante («¡Falta algo!») que equivale a un veto de bolsillo al voto positivo del sistema superviviente: el resultado que sale a la luz es la convicción íntima del paciente de estar mirando a un impostor. En lugar de atribuir esa falta de identificación a un sistema perceptivo defectuoso, el agente echa la culpa al mundo de un modo tan extravagante metafísicamente y tan improbable que apenas caben dudas del poder (el poder político, efectivamente) que normalmente tiene sobre todos nosotros ese sistema deteriorado. Cuando esta hambre epistémica concreta queda insatisfecha, nos da tal ataque que echa por la borda las contribuciones de los demás sistemas.
Entre el olvidadizo cangrejo y el extravagante paciente equivocado de Capgras hay casos intermedios. ¿Puede un perro equivocarse de dueño o no reconocerlo? Según Homero, cuando Ulises regresa a Ítaca después de su odisea de veinte años, disfrazado con harapos como un mendigo, le reconoce su antiguo perro Argo, mueve la cola, agacha las orejas y después muere. (Y habría que recordar que Ulises se enjuga secretamente una lágrima). Así como hay razones para que un cangrejo reconozca (o intente reconocer) la identidad de su propio nido también hay razones para que un perro reconozca (o intente reconocer) la identidad de su amo, entre otras muchas cosas importantes de su mundo. Cuanto más fuertes son las razones para reidentificar las cosas, más beneficioso es no cometer errores y por ello mayor inversión se hace en maquinaria perceptiva y cognitiva. Los modos de aprendizaje avanzado dependen, ciertamente, de capacidades preferentes de (re)identificación. Sea un caso sencillo, el de un perro que ve sobrio a Ulises el lunes, el miércoles y el viernes, pero le ve borracho el sábado. De este conjunto de experiencias se pueden sacar diversas conclusiones lógicamente disponibles: que hay hombres borrachos y sobrios, que un hombre puede estar borracho un día y sobrio otro, que Ulises es uno de esos hombres. El perro no podría (lógicamente no podría) saber el segundo o el tercero de los hechos de esta secuencia de experiencias independientes a menos que tuviera algún modo (falible pero fiable) de reidentificar al hombre como ese mismo hombre en una y otra experiencia. (Millikan, 1997). (Podemos ver una aplicación más drástica del mismo principio en el curioso hecho de que no podemos saber [como cuestión de lógica] qué aspecto tenemos mirándonos en un espejo a menos que tengamos algún otro medio de saber que la cara que vemos es la nuestra. Sin esa identificación independiente no podríamos saber más de nuestro aspecto mirándonos en un espejo que si miráramos una fotografía que por casualidad fuera de nuestra cara).
Los perros viven en un mundo conductual mucho más rico y más complejo que el mundo del cangrejo, con más oportunidades de subterfugios, de faroleo y de disfraces, y por ello tienen mayores beneficios que obtener del rechazo de pistas equívocas. Pero además, los sistemas de un perro no tienen por qué estar a prueba de estupideces. Si el perro mete la pata en la identificación (del tipo que sea) podemos caracterizarlo como un caso de identidad equivocada sin haber llegado a la conclusión de si el perro es capaz de pensar la proposición del comportamiento como si creyera en ella. El comportamiento de Argo en el relato homérico es conmovedor pero no podemos consentir que el sentimentalismo nuble nuestras teorías. Argo también podría amar los aromas del otoño y responder con alegría todos los años cuando le llegara al morro la primera bocanada de los frutos maduros, pero ello no querría decir que tuviera manera alguna de distinguir entre tipos de estaciones recurrentes, como el otoño, e individuos que regresan, como Ulises. Para Argo ¿Ulises no es nada más que una colección organizada y agradable de olores y sonidos, imágenes y sentimientos, una especie de estación recurrente pero irregular (¡llevaba veinte años sin volver!) durante la cual se ven favorecidos determinados comportamientos? Es una estación que generalmente está sobria aunque a veces se sabe también que está ebria. Nosotros podemos ver, desde nuestra peculiar perspectiva humana, que el éxito de Argo en este mundo dependerá generalmente de lo mucho que su conducta se aproxime al comportamiento de un agente que, como nosotros los seres humanos adultos, distinga claramente entre distintos individuos. De manera que cuando interpretamos su conducta desde el enfoque intencional, hacemos bien en atribuir a Argo creencias que distinguen a Ulises de otras personas, perros rivales fuertes de otros rivales más débiles, corderos de otros animales, Ítaca de otros lugares, y así sucesivamente. Pero debemos estar preparados para descubrir que esta comprensión aparente del perro presenta algunas lagunas sorprendentes, lagunas inconcebibles en un ser humano con nuestro esquema conceptual y por ende absolutamente inexpresables en los términos del lenguaje humano.
Durante milenios han sido muy corrientes los relatos de inteligencia de las mascotas. El antiguo filósofo estoico Crisipo contaba de un perro que podía realizar la siguiente hazaña racional: al llegar a una encrucijada con tres caminos, olía los caminos A y B y sin olfatear el C, corría por éste habiendo razonado que como no hay rastro en A ni en B, la presa debería haber ido por C. Las personas son, en cambio, menos aficionadas a contar las estupideces de sus mascotas y suelen resistirse a las conclusiones que les marcan las lagunas que descubren en sus habilidades. Un perrito tan listo y sin embargo ¿no es capaz de averiguar cómo desenrollar la correa mientras gira en torno a un árbol o a una farola? Y no se trata, como podría parecer, de una prueba de inteligencia desleal para un perro, comparada, por ejemplo, con un examen de sensibilidad hacia la ironía en la poesía, o de la apreciación de la transitividad de la expresión comparativa más caliente que (si A está más caliente que B y B está más caliente que C, ¿entonces A está más caliente o más frío que C?). Pero son pocos los perros que pueden superarla, si es que la supera alguno. Y con toda su inteligencia, los delfines son extrañamente incapaces de averiguar que fácilmente podrían saltar hacia su libertad por encima de las redes atuneras. Saltar fuera del agua no es una acción antinatural para ellos, lo que hace más llamativa su cerrazón. Como los investigadores van descubriendo con regularidad, cuanto más ingeniosamente se investigan las capacidades de los animales no humanos, más probable es descubrir abruptas lagunas en ellos. La capacidad de los animales para generalizar a partir de la explotación de sus conocimientos es muy limitada. (Véase Cheney y Seyfarth, How Monkeys See the World [Cómo ven los monos el mundo], 1990 para un informe esclarecedor de esta pauta de investigaciones en las mentes de los monos tumblu[7]).
Nosotros, seres humanos, gracias a la perspectiva que obtenemos de nuestra capacidad de reflexionar según nuestra peculiar manera, podemos discernir los fallos de rastreo que quedarían muy fuera del alcance de otros seres. Supongamos que Tom ha llevado encima, durante muchos años, una moneda de la suerte. Aunque no la llame de ninguna manera en especial nosotros la llamaremos Amy. Tom se lleva a Amy a España, la pone en la mesilla de noche al dormir, y así sucesivamente. Un día, en un viaje a la ciudad de Nueva York, cediendo a un impulso, Tom lanza a Amy a una fuente donde se mezcla con las demás monedas de un penique, absolutamente indistinguible, tanto para Tom como para nosotros, de todas ellas… o por lo menos de todas aquellas que llevan la misma fecha de acuñación que Amy. Y aun así, Tom puede reflexionar sobre este desarrollo. Puede reconocer la verdad de la proposición de que uno, y sólo uno, de esos peniques es su moneda de la suerte que siempre ha llevado consigo. Puede preocuparse (o sencillamente divertirse) ante el hecho de que ha perdido irremisiblemente la pista de algo con lo que ha mantenido el contacto durante años, por un medio u otro. Supongamos que coja de la fuente una moneda que pueda ser Amy. Puede darse cuenta de que una de las dos proposiciones siguientes, y exactamente una de las dos, es verdadera:
- El penique que tengo ahora en la mano es el penique que traje a Nueva York.
- El penique que tengo ahora en la mano no es el penique que traje a Nueva York.
No hace falta ser un científico espacial para darse cuenta de que una de las dos tiene que ser verdad, aunque ni Tom ni nadie más en la historia del mundo, del pasado o del futuro, sepa decir cuál. Esta capacidad que nosotros poseemos de montar hipótesis sobre la identidad, e incluso de comprobar dichas hipótesis en la mayoría de las circunstancias, es bastante ajena a todas las demás criaturas. Las prácticas y proyectos de muchas criaturas les exigen rastrear y reidentificar individuos (sus madres, sus parejas, sus presas, sus superiores y sus subordinados en el grupo) pero no hay prueba que parezca indicar que se den cuenta de que cuando hacen eso, es eso lo que están haciendo. Su intencionalidad nunca llega al punto de la concreción metafísica a la que puede elevarse la nuestra.
¿Cómo lo conseguimos nosotros? No hace falta ser un científico espacial para tener semejantes pensamientos, pero lo que sí hace falta es una criatura gregoriana que entre sus herramientas mentales cuente con el lenguaje. Pero para poder utilizar el lenguaje tenemos que estar equipados con los talentos que nos permitan extraer estas herramientas mentales del medio (social) en que residen.