VII. FANTASMAS DEL INFIERNO
Cuando, en 1784, se produjo el descubrimiento del primer pterodáctilo en la piedra caliza de la localidad bávara de Eichstätt, hecho por el naturalista italiano Cosmo Alessandro Collini, el hallazgo constituyó el último enigma para todos los curiosos científicos de Europa. El pterodáctilo sería, junto con el mosasaurio, el fósil que probaría de forma dramática la realidad de la extinción. Además, dejó bastante manifiestas las grandes desigualdades que existen entre las formas actuales de vida y las del lejano pasado. Cuvier, que sería el que reuniría las pruebas fósiles que harían creíble la extinción, tenía graves dudas acerca de las interpretaciones contemporáneas de este extraño animal prehistórico. El fósil fue llevado al Grand Ducal Museum, en Mannheim, del cual Collini era el director. Collini daba por sentado que la criatura era un habitante de los antiguos mares, pero los doctores alemanes que visitaron Mannheim tenían otras ideas. S. T. Sömmering sostenía que se trataba de una criatura intermedia entre las aves y los murciélagos, mientras que Lorenz Oken no se podía decidir entre un mamífero y un reptil. Cuvier se dio cuenta del parecido superficial con los murciélagos y los vampiros en su forma general, su pico era alargado como el de las chochas pero provisto de dientes como el hocico de los cocodrilos, sus vértebras y las patas eran similares a las de un lagarto, sus dedos en forma de garras eran como los de los murciélagos y su cuerpo estaba cubierto de escamas como en las iguanas. Cuvier era considerado como uno de los paleontólogos más perspicaces de Europa, reputación que se había ganado gracias a su brillante técnica de anatomía comparada. Por consiguiente, era de él de quien se esperaba con impaciencia la solución del enigma. Después de haber examinado el fósil de Collini, a principios del siglo XIX, prestando una especial atención a su cráneo y articulación de los maxilares, Cuvier afirmó que, con toda seguridad, se trataba de un saurio, un reptil del antiguo mundo. Sus cuatro dedos alargados sostenían las alas que lo capacitaba para volar y, por esta razón, lo llamó Pterodactylus o «dedo de ala». Aunque pertenecía a la clase de los saurios y, por lo tanto, estaba emparentado con los reptiles modernos, Cuvier hizo la siguiente advertencia: de todos los seres del mundo antiguo, éste era el más extraordinario y, si se le devolviera la vida, no se parecería a ningún otro tipo de forma de vida[1].
Ya no se podía dudar durante más tiempo que razas enteras de animales habían desaparecido de la faz de la tierra, o que la vida primitiva había tenido un aspecto arcaico. A pesar de todo, la idea de que el pterodáctilo había sido una criatura marina estaba tan arraigada que muchos se opusieron a Cuvier. Los pterodáctilos traerían muchos problemas en el futuro. «En Inglaterra —dijo Harry Govier Seeley más tarde—, son clasificados como reptiles, principalmente debido a la influencia que tuvo sobre ellos la disertación del barón Cuvier[2]». Sin embargo, los doctores alemanes no aceptaron las ideas de Cuvier, simplemente, debido a la gran reputación de la que gozaba el científico francés. En 1830, Johannes Wagler vio en el pterodáctilo un parentesco con los ictiosaurios y los plesiosaurios, que habitaban los océanos del mesozoico. Al igual que éstos, nunca abandonó el agua «pero nadaba en la superficie como un cisne y buscaba los alimentos en el fondo del mar». Se imaginaba que los largos brazos funcionaban como las aletas de los pingüinos o de las tortugas, y que los tres pequeños dedos en forma de garras servían para agarrar a «las hembras en el proceso reproductor[3]». Al año siguiente, 1831, Georg August Goldfuss, profesor de zoología en la Universidad de Bonn y una eminente autoridad alemana en animales fósiles, confirmó el diagnóstico de Cuvier: los pterodáctilos eran reptiles. Sin embargo, añadió Goldfuss, se trataba de reptiles de aspecto muy parecido a las aves y, probablemente a consecuencia de esta creencia, dijo haber observado crestas parecidas a las plumas grabadas en el fósil, adorno que fue rechazado con vehemencia por sus contemporáneos.
Durante la década de 1820, muchos de los recolectores ingleses de fósiles creyeron que los delgados huesos de sus colecciones no pertenecían a aves, sino que eran restos de esta extraña criatura reptil descrita por Cuvier. Pero no fue hasta diciembre de 1828 cuando se descubrió un esqueleto casi completo. Prácticamente, todos los hallazgos de los primitivos reptiles dominantes en Inglaterra fueron hechos por aficionados, y muchos de éstos por uno en particular, una joven y talentosa dama llamada Mary Anning. Los contemporáneos menos dotados tienen la tendencia de otorgar a las personas con talento alguna desgracia ocurrida en su juventud que justifique este don anormal, y Mary Anning no fue ninguna excepción. A la tierna edad de un año, ella y su niñera fueron alcanzadas por un rayo. La niñera falleció pero la niña «pudo ser reanimada metiéndola en un baño de agua caliente; no había sufrido lesión alguna. Antes había sido una niña inactiva, pero después de este accidente, se convirtió en una muchacha más vivaracha e inteligente[4]». Habiéndose dado cuenta de su talento, abrió una tienda con su hermano en Lyme Regis, Dorset, y allí vendía los fósiles que recogía durante sus paseos diarios a lo largo de los vecinos acantilados. En 1811, cuando sólo contaba 11 años de edad, halló un esqueleto de ictiosaurio y, en 1824, el primer plesiosaurio completo. Todos los geólogos de aquella época le ofrecieron su amistad y muchos fueron a Lyme Regis, entonces un lugar de moda, a pasar sus vacaciones para poder pasear con Mary por los acantilados. Cuando, en 1828, desenterró el primer pterodáctilo inglés se lo mostró a un frecuente visitante suyo, el reverendo William Buckland de la Universidad de Oxford. Buckland le describió el fósil a la Sociedad Geológica de Londres al año siguiente (aunque la descripción no fue publicada hasta 1835, una ilustración gráfica de la lentitud de la ciencia a principios del siglo XIX; el ritmo sería acelerado considerablemente cuando Marsh y Cope comenzaron a disputarse la prioridad). Aunque ya había sido preparado por Cuvier para ver algo asombroso, la total sorpresa que se llevó Buckland cuando le fue entregada la ilustración hizo que repitiera las palabras de Cuvier. Era, dijo, «un monstruo que no se parecía a nada jamás visto o conocido en la tierra, exceptuando los dragones de las novelas o libros de caballerías[5]». La técnica anatómica de Cuvier, tan inspiradora para Buckland, había revelado al animal como una criatura voladora. Sin embargo, Buckland estaba claramente influido por algunas de las ideas rivales y volvieron a aparecer viejas confusiones. Buckland estaba desconcertado por los cortos dedos en forma de garras: debieron de ser fuertes garras, pensó, que permitirían al animal reptar, o trepar o colgarse de los árboles. ¡Al año siguiente, en su contribución a los Bridgewater Treatises on the Power Wisdom and Goodness of God as manifested in the Creation, agotó todas las posibilidades de hacer de él también una criatura acuática! «Así, como el demonio de Milton, cualificado para todos los servicios y todos los elementos, la criatura era un compañero idóneo para los emparentados reptiles que pululaban en los mares o se arrastraban en las orillas de un turbulento planeta:
El demonio,
a través de los pantanos, cuestas empinadas, estrechos, terrenos quebrados y vegetación densa o escasa, sigue su camino con la cabeza, manos, alas o pies, y nada o se sumerge, o vadea, o se arrastra o vuela.
EL PARAÍSO PERDIDO, Libro II
»Con bandadas de criaturas de este tipo, volando en el aire, y multitudes de ictiosaurios y plesiosaurios no menos monstruosos en los mares, y cocodrilos gigantes y tortugas arrastrándose por las orillas de los primitivos lagos y ríos, el aire, el mar y la tierra debieron de estar habitados de forma extraña en los primeros períodos de nuestro joven mundo[6]».
La imagen evocada por Buckland fue reflejada de forma ilustrativa por otro aficionado bastante excéntrico de Dorset, sólo cuatro años más tarde. Thomas Hawkins, un coleccionista de fósiles saurios marinos, reprodujo de forma gráfica la confusión contemporánea e hizo una interpretación teológica que resultaba bastante anacrónica, incluso en 1840. La portada del libro de Hawkins, Book of the Great Sea-Dragons, inchthyosauri and Plesosiosauri, Gedolim Taninim of Moses, Extinct Monsters of the Ancient Earth, había sido confeccionada por John Martin y representaba la carnicería que se producía cuando estos monstruosos seres se juntaban en el mundo preadanita. Los grabados de John Martin también adornaban la obra de Gideon Mantell, Wonders of Geology, donde ilustraba a un Megalosaurus y un Iguanodon luchando encarnizadamente. Estos grabados sólo recibieron elogios de Hawkins y Mantell a pesar de que, como reconstrucciones científicas, aun siendo las primeras de este tipo, más bien parecían caricaturas de apariciones grotescas. Othenio Abel, profesor de paleobiología en la Universidad de Viena, a principios de este siglo, y experto en reconstrucciones, las censuró por ser tan atroces como la fantasía de Julio Verne. Nos recuerdan, dijo sarcásticamente, una «figura fantasmal de uno de los infiernos de Breughel[7]».
Hawkins era un recolector infatigable y, en su afán de «sacar a los antiguos saurios de sus tumbas de roca», tal como los describió Richard Owen, acabó llevándose yacimientos enteros. Mediante este método de demolición general, recogió algunos de los mejores saurios marinos jamás hallados que, con el tiempo, pasaron a ser propiedad del Museo Británico donde aún están expuestos hoy en día. Afortunadamente, los monstruos que habitaban inofensivamente la tierra sólo eran «efigies de razas extinguidas», razas que «eran ajenas a las actuales como lo son, probablemente, las de Saturno[8]». Eran una creación antigua de Jehová, que hacía mucho tiempo ya había abandonado el proyecto. «Perpetúan un diseño que ya no se usa», dijo Hawkins, de forma que sería imprudente intentar emparentar las formas pasadas y presentes. Se trataba de mundos, o al menos reinos, diferentes. ¡Estos gigantes marinos ni siquiera pertenecían al reino animal y Hawkins, de acuerdo con esto, creó un nuevo reino bíblico para ellos, el Gedolim Taninim!
Cuando, finalmente, se impusieron las ideas de Cuvier y se determinó definitivamente la forma de locomoción de los pterodáctilos, se disipó la confusión. Antes de que Richard Owen mandara reconstruirlos en el Crystal Palace, en 1854, ya habían sido reconocidos por todos como criaturas voladoras. Los representó posados majestuosamente en la cima de un risco a punto de elevarse en el aire, uno con las alas plegadas y el otro con las alas extendidas. Después de haber llegado a la opinión unánime de que se trataba de voladores, surgió un nuevo problema. Debido a que tanto los pterodáctilos como las aves se habían originado a partir de arcosaurios ancestrales, y ambos tenían alas, algunos pensaron que podrían estar directamente emparentados. Owen rehusó cautelosamente equiparar los pterodáctilos a las aves, a diferencia de St. George Mivart que, como apasionado evolucionista, deseaba hacer de los pterodáctilos los antepasados inmediatos de las aves voladoras. Los pterodáctilos habían aparecido en rocas de épocas mucho más antiguas, observó como prueba de su teoría, y los largos picos y forma general recordaban a las aves. La realización se hizo aún más verosímil cuando los paleontólogos empezaron a obtener pruebas fragmentarias del cerebro del pterodáctilo, del cual se había sospechado desde hacía tiempo que era más grande que el de los reptiles modernos.

q. Ilustración contemporánea de los pterodáctilos realizada por Owen y Hawkins en el Crystal Palace. Hawkins se imaginaba a los reptiles voladores como prototipos del mítico dragón y los construyó de acuerdo con esta idea.
Los esqueletos fósiles bien conservados de pterodáctilos que revelen detalles del cerebro son bastante raros aunque, a lo largo del siglo XIX, se fueron reuniendo pruebas a partir de diferentes fragmentos. Posteriormente, en la década de 1880, el reverendo D. W. Purdon encontró un esqueleto de aspecto prometedor en el esquisto de alumbre del jurásico inferior en Whitby, en Yorkshire, que envió al Geological Survey, en Londres, para su examen. En el Survey, en 1888, E. T. Newton llamó a la criatura Scaphognathus purdoni, en referencia a su descubridor, y, aprovechándose de su estado no fragmentado, sometió el esqueleto a un detallado estudio[9]. Una fractura en el cráneo permitió estudiar la cavidad craneana y sugirió que se había conservado en las rocas un molde del cerebro durante 170 millones de años. Newton, sacrificando los huesos craneanos superficiales, fue recompensado por una impresión craneana parcialmente impecable y, a partir de ésta, confeccionó una réplica del cerebro del «pterosaurio». (El término colectivo de «pterodáctilo» ya estaba siendo abandonado pues sólo se refería a uno de los grupos principales de reptiles voladores; el conjunto del grupo se denomina ahora pterosaurios).
El diminuto cerebro de los reptiles modernos nunca llena por completo la cavidad ósea del cráneo. Por esta razón, los moldes tomados del interior de la caja craneana no son realmente representativos del cerebro que siempre es más pequeño. En las aves y los mamíferos, en cambio, el cerebro muy expandido ejerce presión contra las paredes del cráneo de forma que los moldes internos proporcionan un modelo exacto del tamaño, y tanto del tamaño como de la forma. Newton, que por primera vez examinó detenidamente el cerebro «reptil» de un pterosaurio, estaba sorprendido. «Es indudable que la forma del molde —escribió— representa la forma del cerebro del pterodáctilo, al igual que lo haría un molde tomado del cráneo de un ave o un mamífero; y estos moldes muestran la forma externa natural del cerebro incluso mejor que el mismo cerebro[10]». El cerebro del pterodáctilo también había llenado completamente su caja craneana.
Esto no era el único parecido. El cerebro se divide en tres regiones principales. El cerebelo, situado en la parte posterior del primitivo cerebro vertebrado, está relacionado con el movimiento y el equilibrio y es un área poco desarrollado en los reptiles. Los lóbulos ópticos del mesencéfalo coordinan la visión y, una vez más, son más pequeños en los reptiles y dinosaurios que en los mamíferos y las aves. Lo mismo es válido para los hemisferios cerebrales en el prosencéfalo, que albergan los centros de coordinación superior (una excepción de esto, tal como hemos visto, son los ágiles y listos dromeosaurios y dinosaurios avestruces). El cerebro posterior, o cerebelo, obviamente tiene una gran importancia en las aves ya que controla y coordina las complicadas maniobras necesarias para el vuelo (y también para posarse en algún lugar ya que requiere un sentido del equilibrio mucho más desarrollado que el necesario para mantenerse sobre cuatro patas). El cerebelo se ha desarrollado tanto en las aves, que los lóbulos ópticos han sido desplazados hacia un lado mientras que el dilatado prosencéfalo ha crecido hacia atrás. Los lóbulos ópticos agrandados han tenido que expandirse lateral y ventralmente hacia la base propia del cerebro.
Esto es totalmente desconocido en los reptiles normales y, a pesar de todo, Newton encontró exactamente la misma condición en el Scaphognathus. El cerebro posterior estaba aumentando de tamaño y había adquirido un aspecto bastante avícola, al igual que el cerebro anterior. Estos dos se habían encontrado y excluido a los lóbulos ópticos de la superficie dorsal del cerebro. Los lóbulos ópticos eran grandes pero habían sido desplazados hacia los lados del cerebro. La completa reducción del bulbo olfatorio demuestra que los pterosaurios, al igual que las aves, prácticamente habían perdido su sentido del olfato y que ambos dependían, en lugar de ello, de una vista excelente. En conjunto, las semejanzas entre el cerebro del reptil volador y el de un ave son notables. Los pterosaurios tenían, con mucho, el cerebro más grande (hablando en términos relativos) que cualquier otro reptil y, por lo que se refiere a la inteligencia, habrían hecho que sus primos reptiles parecieran bastante estúpidos, aunque entre los dinosaurios podrían ser equiparados a los dromeosaurios y a los dinosaurios avestruz. Los pterosaurios, supuestamente reptiles inferiores, habían desarrollado de forma independiente un cerebro casi tan eficaz como el de las aves. «Por sí solo, —dijo Seeley en 1901—, el cerebro con forma avícola en un animal sería una prueba, tan válida como cualquier otra, de que su grado de organización era el de una ave[11]».
La razón de la semejanza de cerebros es fácil de comprender. Los pterosaurios tenían tamaños muy variables, que iban desde el diminuto «dragón alado», Ptenodracon, de finales del Jurásico, tan pequeño como un gorrión, hasta los gigantescos Pteranodon y criaturas aún más grandes de finales del cretáceo. La mayoría de los pterosaurios pequeños tenían las alas cortas y anchas que caracterizan a las aves que baten las alas (en contraposición con las aves planeadoras), de forma que es sensato suponer que ellos también batían las alas. (Las alas largas y delgadas, como las de los albatros, de los pterosaurios que aparecieron posteriormente indican hábitos planeadores). Debido a que los pterosaurios adoptaron el mismo tipo de existencia aérea que las aves, debieron necesitar las mismas áreas de coordinación en el cerebro para controlar el equilibrio y poder realizar maniobras complejas. Al igual que en las aves, su vista era obviamente excelente (esto también lo sabemos en base al gran tamaño de las órbitas) y es posible que incluso tuvieran una visión en color.
La idea de que los pterosaurios tenían cerebros como las aves fue confirmada completamente cuando, en 1941, fue examinado otro excelente espécimen. El pterosaurio de Newton había perdido parte del cerebro, pero el espécimen descrito por el eminente neuropaleontólogo Tilly Edinger pudo ser reconstruido completamente[12]. El nuevo espécimen, un diminuto Pterodactylus de sólo unos cuantos centímetros de longitud, y perfectamente conservado en su bloque de Solnhofen, había sido donado al Museo de Zoología Comparada de la Universidad de Harvard por su descubridor, Louis Agassiz, en el siglo XIX. Un gran «lóbulo temporal» en el cerebro anterior le proporcionaba a la criatura un aspecto con cabeza oviforme. En vida, probablemente incluso había tenido un aspecto inteligente. Edinger también quedó sorprendido por la forma de cómo el cerebro posterior y el cerebro anterior habían crecido hasta juntarse como en las aves. Obviamente, observó, «éste es uno de los caracteres que diferencia a todos los pterosaurios de otros reptiles». Cualquier duda que aún pudiera quedar en relación con la inteligencia de los pterosaurios (Newton no quiso destacar demasiado esta característica para no apoyar la teoría de que los pterosaurios evolucionaron hacia las aves), fue rápidamente disipada por Edinger. El Scaphognathus de Newton tenía un pico extraordinariamente largo que hacía que el cerebro pareciese más pequeño en relación con la longitud del cráneo. En otros especímenes, en cambio, las proporciones eran como las de las aves cantoras. Cuando los pterosaurios evolucionaron, sus cerebros se hicieron aún más parecidos a los de las aves, de forma que, en el Pteranodon, la anchura del cerebro anterior era incluso mayor que su longitud al igual que sucede en las aves. Los pterosaurios incluso poseían surcos cerebrales en exactamente las mismas posiciones que en los cerebros de las aves, mientras que los lóbulos ópticos de los pterosaurios que aparecieron posteriormente como el Pteranodon eran similares a los de las aves más avanzadas. En los reptiles, la parte posterior del cráneo está modelada de acuerdo con su necesidad de proporcionar áreas de adhesión para los músculos, mientras que, en los pterosaurios, aves y mamíferos, el cerebro posterior se ha agrandado tanto que ha formado una cúpula cerebelosa que determina la forma de la parte trasera del cráneo. «Por lo que se refiere a la longitud —concluyó Tilly Edinger—, los cerebelos de los pterosaurios se hallan en el límite superior de variación reptil y en el límite inferior de variación avícola», aunque su aspecto fuera puramente avícola. ¡El pterosaurio debió de ser un reptil tan extraordinario como lo es un ave o un mamífero! Sin embargo, aún veremos cuán extraordinario era.
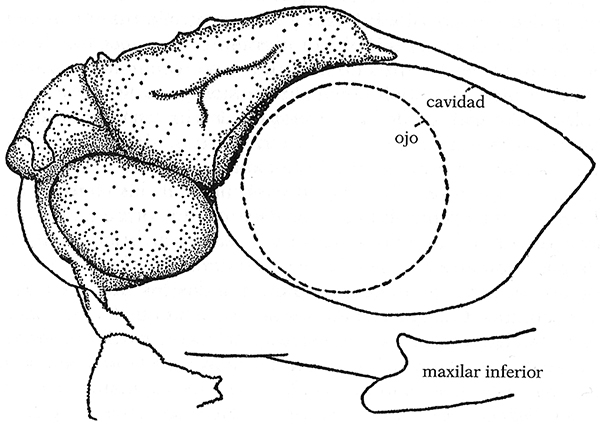
r. Cerebro agrandado del Pterodactylus que hacía presión contra su caja craneana.
Debido a que el funcionamiento de un avión se equipara a sus controles, podemos, suponer, que los pterosaurios eran voladores muy maniobrables. Los mecanismos de control estaban tan sumamente bien desarrollados para el vuelo complejo que, durante algún tiempo, los pterosaurios mantuvieron los suyos en competencia con las aves en vías de desarrollo. La extremadamente activa existencia aleteadora de las aves requiere una gran cantidad de energía. El delicado equilibrio, la compleja coordinación y la pura fuerza requeridos, significan que los músculos deben recibir constantemente oxígeno para que puedan oxidar rápidamente los azúcares, a fin de liberar un flujo continuo de energía. Estas reacciones químicas que producen energía requieren una temperatura alta, constante para una liberación óptima y rápida de la energía en todo momento. Por lo tanto, no es una sorpresa que las aves de sangre caliente, con un elevado índice metabólico, tengan temperaturas corporales medias uno o dos grados más elevadas que en el hombre. Al disponer de un metabolismo rápido, las aves son capaces de permanecer activas durante largos períodos de tiempo. Han hecho todo lo posible para desarrollar estructuras nuevas que permitan que los pulmones puedan absorber la máxima cantidad de oxígeno: oxígeno requerido tanto para mantener una elevada temperatura como para producir la energía necesaria para volar.
La mecánica respiratoria de los mamíferos (una bomba constituida por las costillas-diafragma-pulmones), proporciona una solución al problema de la energía. Sin embargo, las aves emplean un sistema distinto, igualmente eficaz. Aquí, los pulmones son más rígidos pero conducen a extensos sacos aéreos que se ramifican por toda la cavidad corporal. Frecuentemente, existen diez de estos sacos de aire aunque éstos también van formando sacos más pequeños, en forma de vejiga, que se distribuyen por los brazos y las piernas. Los huesos de muchas aves son huecos y tienen pequeñas aberturas específicamente para permitir la entrada de los sacos aéreos (que revisten el interior de los huesos). Los huesos de las extremidades, columna vertebral, etc., son literalmente estructuras llenas de pulmón. Se cree que estos sacos, en forma de globo, actúan como depósitos de aire para el tórax, como un fuelle, bombeados mediante el esternón y el batido de las alas (durante el vuelo, las palomas sincronizan la respiración y el batido de las alas). El bombeo impulsa el flujo de aire a través de los diminutos tubos pulmonares, donde se produce la verdadera absorción de oxígeno y eliminación del dióxido de carbono residual. Y, lo que es de suma importancia, este flujo de aire a través del cuerpo facilita el enfriamiento. Frecuentemente, se puede ver a las aves «jadear» después de haber hecho ejercicio.
Es razonable pensar que los pequeños pterosaurios batían las alas, despegando activamente. En este caso, probablemente necesitaban un rápido intercambio de oxígeno-dióxido de carbono para obtener una producción de energía constante. Al igual que las aves, necesitarían eliminar rápidamente el exceso de calor. Debido a que los pterosaurios poseían pequeños orificios en sus huesos huecos, probablemente también tenían sacos pulmonares, y una bomba de los sacos aéreos. En 1837, Hermann von Meyer utilizó estas aberturas como prueba de que los pterosaurios eran voladores. Los huesos neumáticos de los pterosaurios parecen huesos avícolas llenos de aire, excepto por el hecho de que presentan una envoltura ósea aún más delgada. Los finos huesos de las aves parecen macizos en comparación. Los huesos de los pterosaurios contenían sacos más grandes con más aire. Quizá por esta razón las aberturas, colocadas de forma similar en las aves y en los pterosaurios, siempre eran más grandes en estos últimos.
Seeley dedicó toda su vida al estudio de los pterosaurios mesozoicos y, en 1901, resumió los hallazgos en el libro Dragons of the Air. Aparte de una obra especializada anterior suya, éste era el primer libro dedicado exclusivamente a estas extraordinarias bestias[13]. En éste, estaba poco dispuesto a admitir que los pterosaurios eran reptiles. El cerebro era como el de las aves, prueba del elevado nivel de inteligencia necesario para el vuelo aleteante de un vertebrado. El sistema respiratorio probablemente era similar al de las aves, demostrando una avidez de aire que revelaba una aceleración metabólica. Naturalmente, la conclusión de Seeley fue que los pterosaurios eran de sangre caliente. De otra manera, es difícil imaginárselos volando durante períodos más largos sin producir un demoledor déficit de oxígeno. Un lagarto aleteante, aún estando equipado con la musculatura y coordinación mental necesarias, habría caído prematuramente a tierra por agotamiento. Es especialmente poco probable que los pterosaurios malgastaran la ventaja de tener una ventilación avícola, no modificando el sistema circulatorio. El corazón de los lagartos, con tres cámaras, mezcla la sangre arterial rica en oxígeno con la sangre venosa pobre en oxígeno antes de bombearla al cuerpo. Los cocodrilos, que se originaron a partir de los ancestrales arcosaurios aproximadamente al mismo tiempo que los pterosaurios, tienen un corazón que funcionalmente posee cuatro cámaras. Probablemente los dinosaurios, que se originaron a partir de un mismo tronco, también desarrollaron este tipo de corazón. Éste está presente en los descendientes avícolas de los dinosaurios. Como observó Seeley: «El tipo de corazón que siempre se asocia a estructuras vitales, como las que se pensaba tenía el pterodáctilo, habiendo sido éstas deducidas a partir del molde cerebral y de los orificios neumáticos en los huesos, es el corazón de cuatro cámaras de las aves y de los mamíferos[14]». El sistema circulatorio de los pterosaurios no presentaba ningún rasgo inconfundiblemente reptil, concluyó Seeley.
Sin duda, Seeley tenía razón. Unos eficaces pulmones avícolas proporcionaban el oxígeno y eliminaban el calor y el dióxido de carbono residual necesarios para los vuelos largos. Probablemente, un corazón de cuatro cámaras garantizaba el suministro constante de sangre saturada de oxígeno a los tejidos. Un vuelo sostenido implica una aceleración metabólica. Los pterosaurios eran de sangre caliente como los mamíferos y las aves.
Debido a que los pterosaurios eran endotermos, y muchos de ellos eran pequeños (el Ptenodracon sólo tenía el tamaño de un gorrión), debieron de estar protegidos por un abrigo de piel o plumas, o quizás incluso disponían de algún tipo de aislamiento que no ha podido sobrevivir hasta nuestros días. Una criatura de sangre caliente del tamaño de un ave no habría resistido nunca la pérdida de calor, si no hubiese dispuesto de plumas. No existen aves desnudas y tan sólo uno o dos ejemplos de pequeños mamíferos sin pelo (algunos animales que viven en madrigueras como los topos carecen casi completamente de pelo pero están aislados del frío extremo por sus madrigueras). La afirmación de Goldfuss de que se podían divisar mechones de pelo en los pterodáctilos fosilizados, nunca fue tomada muy en serio en Alemania. Esto no es sorprendente, ya que Cuvier había dicho que la bestia era un reptil y los reptiles son escamosos por definición. Hasta el lúcido estudio realizado por Seeley era inconcebible, al pensar que un reptil pudiera ser de sangre caliente. Anteriormente, ya se habían hecho una o dos sugerencias alternativas, pero el tema era abordado con extrema cautela y sin confianza alguna. Las ideas de Goldfuss impresionaron mucho al zoólogo inglés Edward Newman. Sin embargo, suponiendo que fuera cierto que los pterosaurios tuvieran pelo, Newman también tendría que admitir que no podían haber sido reptiles. Cuando, en 1843, Newman escribió su artículo tenía conocimiento del controvertido maxilar de «opossum» de Buckland hallado en la pizarra jurásica de Stonesfield treinta años antes (de hecho fue el primer mamífero mesozoico en ser descubierto). De forma que se aventuró a sugerir que los pterodáctilos contemporáneos también habían sido marsupiales, sólo que murciélagos marsupiales. Estando expuesto a la mofa pública, insistió en que los anatomistas no adoptaran una actitud reverencial hacia las autoridades: Cuvier y Buckland podrían estar equivocados. «Ahora creo que es posible —dijo Newman cautelosamente— que Cuvier y Buckland no tuvieran razón. Confieso que esto es muy improbable, pero sostengo que es posible. Sea cual sea el concepto que tengamos de ellos, existe el factor humano que nos induce a pensar que ellos también son capaces de cometer errores[15]». La herejía de Newman fue contestada con un silencio torrencial, destino que acontece demasiadas veces a las especulaciones iconoclásticas.
Sólo cuando Seeley comenzó a echar las bases para el cambio en la opinión respecto a la condición del pterosaurio, la cuestión del aislamiento se hizo crucial. Si, como sugería Seeley, se trataba de una criatura de sangre caliente, ¿no debería haber tenido algún tipo de aislamiento? En 1908, Karl Wanderer, de la Universidad de Munich, examinó los pterosaurios en el Museo de Dresde para ver si él podía ver pelo. Goldfuss afirmaba que su fósil presentaba impresiones de piel con una superficie verrugosa, lo cual interpretó como prueba de la presencia de pelo o plumas en vida. Wanderer lo dudaba. Estos «hoyuelos» distribuidos irregularmente no eran debido a otra cosa que a la naturaleza desmenuzable de la roca[16]. Más tarde, en 1927, uno de los paleontólogos alemanes más respetados, Ferdinand Broili, mostró enérgicamente su disconformidad con Wanderer insistiendo en que efectivamente se trataba de impresiones de mechones de pelo y, para demostrarlo, comparó la piel verrugosa de un murciélago con las impresiones de la piel de los pterosaurios. No cabe duda, exclamó, de que los pterosaurios poseían un abrigo de piel[17]. Sin embargo, el artículo cayó oportunamente en el olvido y, si no fuera por algunos recientes hallazgos espectaculares, probablemente se habría perdido por completo.
En 1970, A. G. Sharov, de la Academia de Ciencias Soviética, estaba recolectando fósiles en los depósitos de lagos del jurásico superior en Karatay, Kazajstán, una provincia del sur de la ex URSS y descubrió un pequeño pterosaurio extraordinariamente bien conservado. Las rocas de esta región que contienen los fósiles, al igual que las de Solnhofen y Eichstätt, son de grano muy fino al haberse formado a partir de los sedimentos de un antiguo lago y, al igual que la piedra caliza de Baviera, registran las impresiones de la piel y otras partes blandas de los animales que quedaron incrustados en ellas. El pterosaurio ruso de Sharov estaba completo, mostrando las impresiones de las membranas de las alas y un abrigo de piel. Todo el cuerpo estaba revestido de pelo largo y espeso; en vida, probablemente era blando y lanudo ya que los pelos y mechones individuales parecen estar curvados e inclinados en el fósil. También había pelo en los dedos y las membranas entre los dedos del pie, aunque aquí más corto, e incluso se puede observar en la misma membrana de un ala. Sharov llamó enigmáticamente su bestia Sordes pilosus, que literalmente significa «abrigo de pieles sucio». Sin embargo, también empleó el equivalente ruso de Sordes que, en esta lengua, tiene el significado alternativo de «demonio» o «espíritu maligno», por lo cual el nombre también podría ser traducido, más sugestivamente, como «demonio peludo[18]».
Este descubrimiento demostró inequívocamente que los pterosaurios eran de sangre caliente. Retrospectivamente, es fácil decir que no podría haber sido de otra forma. Sin embargo, el proceso para llegar a esta conclusión fue largo y penoso, abarcando casi dos siglos. Durante este tiempo, nuestra opinión sobre esta extraña bestia ha sufrido una radical transformación. Con la autoridad de Cuvier, el pterodáctilo fue clasificado como reptil, lo cual hacía imposible la existencia de pelo. (Oficialmente aún sigue siendo clasificado como reptil en todos los textos corrientes). Después de la publicación de los estudios de Seeley a principios del siglo XIX, se fue aceptando cada vez más la idea que un vertebrado volador no podría haberse elevado nunca en los aires, a menos que dispusiera de una fisiología con un metabolismo rápido y con gran producción de energía, al menos equivalente a la de un murciélago. Este tipo de fisiología se acompaña de una elevada temperatura interna, que es mantenida estable para garantizar un funcionamiento bioquímico óptimo. En base a estos antecedentes teóricos, el hallazgo de Sharov no debería haber causado gran sorpresa; ya estábamos prevenidos. Con el fin de evitar la pérdida de calor y el desperdicio de energía, los pterosaurios habían desarrollado pelo de forma paralela a los mamíferos. Al igual que los mamíferos, los pterosaurios probablemente pasaban muchas horas cada día limpiándose el pelo. En los pterosaurios primitivos, los dientes y las garras posiblemente habían servido de peines. Cuando en los tipos que surgieron posteriormente se perdieron los dientes, con el fin de perder peso, y las garras ya no llegaron al tronco, posiblemente el pico asumió esta función.
Debido a que los pterosaurios eran bastante inteligentes y de sangre caliente, ¿cuál era su forma de reproducción? ¿Ponían huevos o daban a luz a crías vivas? Y lo que es aún más importante, ¿existían cuidados posnatales? Incluso los cocodrilos de pequeño cerebro son padres dedicados que desentierran a sus crías que salen de los huevos para transportarlas hasta el agua (en sus maxilares). Incluso irán en su ayuda si las oyen chillar cuando están en manos de algún científico incauto. Apenas cabe menos esperar de la forma de atención paterna de los lejanos primos voladores de los cocodrilos.
Debido a que los pterosaurios habían desarrollado el cuerpo volador más ligero de todos los vertebrados, y se habían tomado mucho trabajo para reducir el peso a gran escala, es improbable que las hembras pudieran haber volado llevando a sus crías cuando éstas hubieran alcanzado una fase muy avanzada de su desarrollo. Si los pterosaurios eran vivíparos, las crías probablemente nacían en estado inmaduro, conclusión que también es sugerida por la pelvis muy pequeña de la madre. O, quizá, ponían huevos al igual que las aves. De cualquier forma, los pterosaurios que salían de sus huevos, debido a que eran criaturas inmaduras y de sangre caliente, debieron de depender de sus padres para que les dieran calor, alimentos y más tarde probablemente un entrenamiento de vuelo. Las hembras, al tener un abrigo de piel y ser una fuente de calor, posiblemente empollaban los huevos hasta que salían las crías (el único caso paralelo en el mundo reptil es el de la hembra pitón de la India, que se enrolla alrededor de sus huevos y se calienta a sí misma enviando espasmos de contracción muscular a lo largo de su cuerpo). Los pterosaurios probablemente permanecían junto a sus crías hasta que éstas podían volar[19]. Esto podría significar que los pterosaurios se apareaban, al igual que las aves, durante una época del año —o incluso de por vida—, de forma que el macho pudiera suministrarle a la hembra, que estaría empollando los huevos, los alimentos almacenados en su bolsa. Los pterosaurios habían ascendido a un nuevo nivel social, caracterizado por la unidad familiar temporal. Puede incluso que hubiera existido una organización colonial más avanzada, en la cual los vulnerables polluelos —especialmente del gigantesco Pteranodon—, formarían manadas y estarían protegidos por uno o dos adultos. (En la arenisca verde de Cambridge, en Inglaterra, se han encontrado miles de fragmentos de pterosaurios más pequeños, sugiriendo que posiblemente habían sido gregarios, pasando la noche juntos en colonias como los murciélagos. Sin embargo, en este caso es más probable que estos huesos fueran arrastrados por el agua quedando amontonados mucho tiempo después de haberse producido la muerte de los individuos vastamente distribuidos[20]).
Seeley concluyó su clásico informe sobre los dragones voladores, u «ornitosaurios» como él los llamaba, con una reafirmación de su teoría de que eran algo más que humildes reptiles: «estos fósiles han demostrado que los ornitosaurios tienen órganos vitales blandos en común con los dinosaurios y las aves», escribió y que habían desarrollado «formas especiales de órganos respiratorios y cerebros que los distinguían de los reptiles existentes[21]». Estas ideas fueron formuladas a principios de este siglo y Seeley, que murió sólo ocho años más tarde, no llegaría a saber nunca que sus ornitosaurios eran peludos, demostrando así fuera de toda duda que eran, tal como él siempre había pensado, de sangre caliente. Su nivel de organización era sumamente alto y, aunque se habían originado a partir del mismo tronco ancestral que los dinosaurios, habían evolucionado más que sus primos. Los dinosaurios también eran de sangre caliente, es cierto, pero los pterosaurios habían desarrollado el pelo como los mamíferos y un cerebro como las aves, de forma que, aun no siendo endotermos más eficientes, con toda seguridad eran más inteligentes. Habían superado la barrera térmica y evolucionado a partir de la clase reptil en la misma medida que las aves o los mamíferos. Bakker y Galton afirmaron, cuando establecieron la clase dinosaurios, que entre los animales vertebrados superiores «la adaptación más importante fue el paso de la endotermia[22]». Esto había ocurrido en los reptiles semejantes a los mamíferos y en los antepasados de los dinosaurios, dando lugar a nuevos niveles de organización y provocando profundos cambios en los tejidos blandos, que permitieron la producción constante de energía y una mayor actividad. Bakker y Galton continuaron: «Probablemente los antepasados dinosaurios de las aves ya eran endotérmicos y tenían un metabolismo de alto rendimiento, características clave que diferencian a los dinosaurios de los cocodrilos y de otros arcosaurios extinguidos». Esto no es del todo verdad, tal como ya se ha comentado anteriormente, ya que los mismos criterios que le permiten a Bakker declarar a los dinosaurios endotermos, probablemente también son atribuibles a los antepasados seudosuquianos de los dinosaurios, en el caso de que fueran de sangre caliente. Finalmente, esta diferencia podría ser ahora ampliada para incluir a los pterosaurios. La endotermia posiblemente diferencie a los dinosaurios de los cocodrilos, y sea una justificación para mantener a estos últimos como reptiles y elevar a los primeros a una nueva clase. Sin embargo, los pterosaurios también habían evolucionado. Se habían vuelto endotermos y, además, tan eficientes como las aves. Parece bastante absurdo dejarlos en la clase de los reptiles y, al mismo tiempo, elevar de clase a los dinosaurios. Seeley no lo hubiese aprobado.
Si los pterosaurios aún viviesen, inmediatamente reconoceríamos que no eran reptiles. Aristóteles —el gran recolector y clasificador de la antigüedad— nunca habría incluido a un animal de sangre caliente, peludo e inteligente, en el grupo de las serpientes. Esta tradición, de no proceder de esta forma, se habría transmitido hasta nuestro siglo XX, tal como ocurre con las aves y los mamíferos. Desafortunadamente, los pterosaurios se extinguieron a finales del cretáceo y sólo a partir de los huesos fósiles pudieron ser revividos en la mente de los hombres; huesos, además, que inicialmente proporcionaron pocas pistas en relación con la fisiología o inteligencia. Estas características han sido determinadas laboriosamente a lo largo de varias décadas, de forma que ahora nos podemos hacer una imagen bastante clara del pterosaurio en vida que se parece muy poco a los reptiles.
Reconocemos que, aunque los mamíferos se originaron a partir de los reptiles, se trata de criaturas distintas. Asimismo, a los pterosaurios, cuya fisiología tan distinta a la de los reptiles es la prueba de que hacía ya mucho tiempo que se habían distanciado de los reptiles, se les debería otorgar la condición de clase en reconocimiento a su elevado nivel de organización.
Ahora disponemos de todas las pruebas y podemos especular acerca del origen y de las subsiguientes radicaciones de los endotermos no mamíferos del planeta mesozoico y, al mismo tiempo, sugerir por qué determinados grupos fueron capaces de «explotar» cuando lo hicieron. En resumen, podemos exponer una teoría unificada que relaciona estas radiaciones de los vertebrados mesozoicos con los logros fisiológicos subyacentes. Probablemente, los seudosuquianos lograron por primera vez una fisiología de sangre caliente a principios del triásico —hace 210 millones de años— y evolucionaron hacia los paradinosaurios, algunos de cuyos miembros más tardíos (por ejemplo, el Ornithosuchus) llegaron a tener un gran parecido con los dinosaurios carnívoros bípedos. Los dinosaurios ornitisquianos y saurisquianos aparecieron aproximadamente a mediados del triásico, explotando al máximo su legado fisiológico y postural seudosuquiano y asumiendo rápidamente una posición dominante en el orden del mundo mesozoico. Sin embargo, otros seudosuquianos bípedos del triásico medio, con brazos libres de la carga de su función de soporte y locomoción, empezaron a planear y desarrollaron una membrana de piel que se extendía entre los brazos y el tronco. Existen pruebas de la existencia de esta fase intermedia, el seudosuquiano planeador Podopteryx (literalmente «pie alado»), encontrado por Sharov en sedimentos del triásico superior en Rusia[23]. (El Podopteryx vivió en una época demasiado tardía como para haber sido el verdadero antepasado de los pterosaurios). Los seudosuquianos estaban «preadaptados» para volar si, en efecto, ya poseían una fisiología de metabolización rápida. De este modo, pudieron elevarse en los aires como criaturas que batían las alas y, a principios del triásico tardío, aparecieron los primeros pterosaurios en los cielos[24]. Es bastante probable que hubiera un único origen de la endotermia entre los «saurios» del mesozoico que se remontaba a los tecodontes de principios del triásico, y que esto desencadenó dos explosiones principales, la de los dinosaurios y la de los pterosaurios. Estos descendientes endotermos seudosuquianos se dieron cuenta del potencial inherente a su legado fisiológico y se convirtieron en unos de los vertebrados terrestres que más éxito han tenido en la historia. (Véase el árbol genealógico de los vertebrados superiores en el apéndice I). La sustitución de un origen único por dos o incluso tres orígenes independientes de la endotermia, simplifica en gran manera nuestra visión de la historia de los vertebrados además de satisfacer la exigencia científica de sencillez. El resultado es una síntesis estéticamente grata que satisface ampliamente, por su amplia envergadura y gran poder aclaratorio (que, al fin y al cabo, es el objetivo final de la explicación científica).
A pesar de que los pterosaurios más pequeños eran aleteadores, a finales del mesozoico se desarrollaron extraños planeadores gigantescos, entre ellos los animales voladores más grandes que han existido en el mundo. Anteriormente a 1840, ninguna especie conocida superaba, en tamaño, a un gran murciélago de la fruta. Esto no era extraño, dijo Richard Owen en aquella época, ya que «a priori era fisiológicamente improbable que la organización de sangre fría de un reptil pudiera ser capaz de elevar en los aires una masa más grande de la que podían levantar los mamíferos de sangre caliente[25]». En 1845, el secretario de la Sociedad Paleontográfica, James Scott Bowerbank, describió un nuevo espécimen de la creta de Kent que presentaba una envergadura de, al menos, 2,44 o 2,74 metros. En lugar de modificar su teoría de que los pterodáctilos eran de sangre fría, Owen exclamó que Dios una vez más estaba demostrando su omnipotencia al haber logrado lo que parecía imposible. Este espécimen, dijo Owen, «demuestra que las manifestaciones de la fuerza del Creador en tiempos pasados sobrepasan cualquier cálculo basado en la naturaleza real[26]». Posteriormente, en 1847, Bowerbank excavó un pterodáctilo que no superaba al otro en longitud, pero que tenía casi el doble de tamaño, con una envergadura que alcanzaba los 4,5 o 4,8 metros de punta a punta y al que llamó Pterodactylus giganteus. Fue una criatura de este tamaño la que Owen mandó reconstruir a escala natural en el Crystal Palace en Syndenham.
Después de la exploración del Oeste, la atención se desvió hacia el Nuevo Mundo. En el otoño de 1870, O. C. Marsh (acompañado de Col. W. F. Cody, más conocido como «Buffalo Bill») dirigió una expedición del Yale College en la región de las Montañas Rocosas en busca de nuevos fósiles y, como siempre, al eminente paleontólogo le aguardaba una gran bonanza. El equipo de Marsh estableció su campamento en la bifurcación norte del río Smokey, al oeste de Kansas, y comenzó a explorar los depósitos de creta del cretáceo superior.
Kansas era el territorio de Cope y la creta había proporcionado una rica variedad de espectaculares criaturas al recién descubierto rival de Marsh, que incluían muchos mosasaurios y el plesiosaurio marino de cuello largo, Elasmosaurus. En realidad, Cope no había visitado Kansas. Los fósiles, de hecho, le habían sido enviados a Filadelfia donde los describió. El Elasmosaurus fue enviado en 1868 y el momento de su llegada a Filadelfia coincidió con el del experto inglés en reconstrucciones, Benjamin Waterhouse Hawkins, que estaba buscando reptiles prehistóricos para su Museo Paleozoico. De esta forma, ocurrió que, apenas Cope había descrito el animal, Hawkins ya había construido un modelo de tamaño natural en el centro de Nueva York. Una vez desencadenada la guerra entre Marsh y Cope (provocada por Cope que no había sido capaz de distinguir entre la cabeza y la cola del Elasmosaurus), comenzó la apasionada carrera en busca de nuevos descubrimientos. Marsh cruzó las Montañas Rocosas por primera vez en 1868 y, pensando que la recolección de huesos en Kansas era buena, volvió con el equipo de Yale en 1870. Marsh inmediatamente desenterró tres nuevas especies de mosasaurios, uno de los cuales alcanzaba una longitud de 9 metros. Durante las excavaciones, se tropezó con algunos huesos problemáticos; éstos eran largos, delgados y, a primera vista, se parecían a los huesos de las patas de las aves zancudas. Al igual que los huesos avícolas eran ligeros, tenían paredes delgadas y eran neumáticos. Los restos de los huesos obviamente procedían de dos especies distintas. Marsh estaba preocupado porque las articulaciones eran completamente diferentes a las de las aves y, a pesar de su gran tamaño, sólo podían ser comparadas con la articulación de los dedos de las alas de los pterosaurios ingleses. Sin embargo, el dedo era desproporcionado; a pesar de que Marsh sólo tenía un diminuto fragmento de un metacarpiano, éste medía 17 centímetros de longitud. «¡Esto indicaría —dijo Marsh proféticamente— una envergadura de las alas de no menos de 6 metros de largo[27]!». La temporada de recolección estaba llegando a su fin, de forma que el entusiasmo tuvo que ser contenido. Marsh se vio obligado a esperar hasta el verano siguiente para poder confirmar sus especulaciones. En el verano siguiente, 1871, el talento de Marsh para descubrir fósiles resultó en un hallazgo que incluso eclipsó parcialmente a sus reptiles alados. Cuando volvió a la creta de la región del río Smokey, en Kansas, descubrió el primer esqueleto de un ave no voladora del cretáceo, el Hesperomis, o «ave del Oeste», en los lechos que contenían los fósiles de pterosaurios[28]. (Los simpatizantes de Cope insisten en que no fue Marsh sino Benjamin Mudge quien, en realidad, encontró los restos del Hesperomis y advirtió su importancia. Mudge estaba a punto de enviárselos a Cope cuando apareció en escena Marsh y convenció a Mudge para que se los entregara). Los enormes pterosaurios obviamente habían vivido junto con aves de aspecto bastante moderno (moderno en comparación con el arcaico Archaeopteryx a pesar de que el Hesperomis, al igual que los dinosaurios-ave, tenía dientes). La profecía de Marsh acerca del gigantesco tamaño de los reptiles voladores se confirmó dramáticamente en este viaje. Encontró nuevos especímenes con los que pudo sumar la longitud de los diversos huesos aislados de alas. El metacarpiano de la mano, del cuál ya había encontrado un pequeño fragmento, medía 40 centímetros mientras que el primero de los cuatro huesos del dedo alargado del ala medía 43 centímetros. El brazo era corto y medía 18 centímetros. Esto hacía un total de más de 1,20 metros sin contar los tres huesos de los dedos de las alas que faltaban. «Las mediciones reseñadas anteriormente de los huesos de las alas indicarían una longitud total de las alas de, al menos, 2,6 metros y una envergadura de ambas alas de 5,5 a 6,1 metros. Por lo tanto, la presente especie incluye a algunos de los “dragones voladores” más grandes jamás descubiertos[29]». Aun hallándose en los primeros días de su carrera en busca de fósiles, Marsh difícilmente podría haber encontrado algo más espectacular para aventajar a su rival. En una posición algo más elevada del mismo yacimiento, desenterró un pterosaurio aún más grande cuyos huesos de las alas eran la mitad más anchas que las del espécimen anterior, que ya había sido bautizado con el nombre de Pterodactilus occidentalis. Marsh, al que obviamente se le acabaron los superlativos, veía al nuevo espécimen como «uno de los pterosaurios más gigantescos. ¡Tenía al menos el doble de volumen que el Pterodactylus occidentalis y probablemente la distancia, medida de punta a punta de las alas completamente extendidas, sería de aproximadamente 6,7 metros!».
Sin duda fue un gran triunfo personal para Marsh. El hallazgo de los primeros Pterosaurios americanos le había correspondido a él, como era justo. Cope, entretanto, no sería dejado fuera. Cuando, al final de la segunda temporada de excavaciones, Marsh abandonó Kansas, Cope inmediatamente se dirigió a los mismos lugares y comenzó a excavar frenéticamente a fin de conseguir su propia colección de valiosos fósiles de la creta. (Cuando, una década más tarde, el Popular Science Monthly publicó un artículo sobre la vida de Cope, éste adelantó la fecha de la época de sus exploraciones al Oeste a 1870 para que coincidiera con la de Marsh). Tan pronto como, hubo marchado Marsh, Cope trabajó con rapidez en un intento de extraer los fósiles y asignarles nombres antes de que lo pudiera hacer Marsh. La prioridad en la asignación del nombre de las bestias era de suma importancia para Cope. El 1 de marzo de 1872 leyó un informe ante la Sociedad Filosófica Americana, describiendo sus hallazgos de pterosaurios. Llamó a los dos saurios gigantes alados Ornithochirus o «pies de ave», en oposición al Pterodactylus de Marsh. Pero llegó demasiado tarde. Tal como señaló con gran placer el portavoz de Marsh —el American Journal of Science publicado en la ciudad natal de Yale, New Haven—, el artículo de Cope fue distribuido en forma impresa el 12 de marzo mientras que la contribución de Marsh había aparecido cinco días antes, el 7 de marzo. Marsh había conservado la prioridad por un pelo.
La creta de Kansas resultó ser un verdadero cementerio de estos enormes saurios voladores y Marsh, seguido más tarde por Willinston, excavaron docenas de especímenes fragmentarios durante las siguientes décadas. Marsh rebautizó a su gigantesca especie con el nombre de Pteranodon, que literalmente significa «alado y sin dientes». Se trataba de un pterosaurio de aspecto extraordinario, con mandíbulas que formaban un largo pico sin dientes y que mantenía el equilibrio con la ayuda de una cresta ósea alargada que se levantaba en la parte trasera del cráneo. La criatura era literalmente toda alas que, probablemente, superaban los 7 metros de envergadura mientras que el tamaño de su diminuto cuerpo no superaba el de un pavo. Un contemporáneo más pequeño de Kansas, el Nyctosaurus, o «lagarto de la noche», ilustra la misma gran desproporción con un cuerpo de 15 centímetros de longitud y alas de 2,4 metros de envergadura.
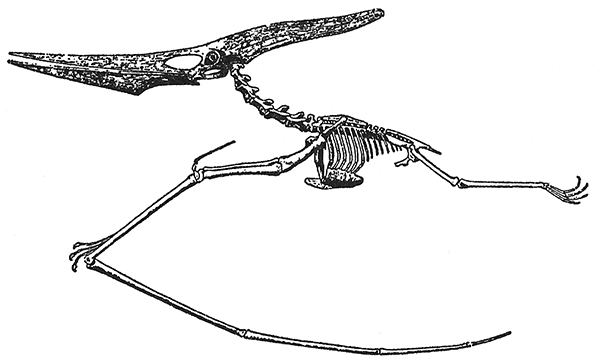
s. El esqueleto del pterosaurio Pteranodon de alta mar. Estaba construido como un planeador artificial. El fuselaje en forma de caja, reforzado, soportaba la tensión de alas de 3,6 metros de longitud. El hueso pteroideo de la muñeca probablemente sostenía una membrana elevadora que llegaba al cuello.
Las largas extremidades traseras colgantes del Pteranodon eran definitivamente inadecuadas para el movimiento terrestre normal y es difícil imaginarse a estas torpes criaturas caminar para el suelo. A diferencia de los murciélagos, que pueden doblar sus alas a lo largo de su cuerpo (e incluso así tienen dificultades para caminar sobre cuatro patas), el cuerpo del Pteranodon no podría de ninguna manera haber sido envuelto por sus largas alas. Debido a que los dedos de las alas sólo podían ser flexionadas en los nudillos, en el punto medio de las alas, los dedos de 1,8 metros de longitud probablemente estarían dirigidos hacia arriba cuando el animal avanzaba a rastras. Cualquier forma de desgarbada locomoción habría resultado extremadamente difícil. Los primeros pterosaurios tenían una articulación entre el hombro y el ala consistente en cóndilos y carillas articulares. Sin embargo, las especies que aparecieron más tarde presentaban una charnela que sólo permitía movimientos ascendentes y descendentes[30]. «Quizás el método de desplazamiento en tierra más factible para ellos —dijeron Hankin y Watson en 1914— era que, una vez se habían posado sobre tierra, cayeran sobre sus estómagos y se fueran arrastrando por el suelo, como hacen los pingüinos, dándose impulso con sus patas traseras y salvando los obstáculos con un ocasional alzamiento de las alas[31]». Naturalmente, en algún momento debían de posarse sobre tierra y, por lo tanto, debían de ser capaces de desplazarse por tierra aunque resulta difícil imaginárselos caminar a modo de pingüinos aunque sólo sea porque poseían unos enormes picos. Quizás empleaban los dedos de las alas para levantar el cuerpo del suelo. Con una locomoción en tierra tan torpe, el Pteranodon habría sido una fácil víctima para cualquier dinosaurio carnívoro que estuviera cerca. Sin embargo, parece más probable que los pterosaurios más grandes volaran casi constantemente, al igual que lo hacen los albatros, y quizá se posaban sobre tierra sólo una vez al año para reproducirse. Las patas traseras, extraordinariamente parecidas a las de los murciélagos, sugieren que el Pteranodon se podía colgar con la cabeza hacia abajo como hacen los murciélagos cuando se hallaba en tierra. Debido a que era demasiado grande como para haber sido un trepador de árboles, se pensaba que descansaba en salientes de las rocas. Sin embargo, probablemente cerca de los lagos del interior de Kansas no había acantilados, por lo cual resulta difícil aceptar la idea popular de que los Pteranodon habían sido habitantes de la costa donde podrían estar seguros[32].
Al igual que los albatros, el Pteranodon probablemente pasaba gran parte de su vida adulta en el aire. Para poder hacer esto, y sin embargo crecer tanto, era necesaria una reducción bastante drástica del peso. Generalmente, no se aprecia suficientemente lo ligero que era en realidad este planeador con alas de 7 metros de envergadura. Debido a esto, se han imaginado todo tipo de ideas fantásticas para explicar cómo el Pteranodon permanecía en vuelo. Se ha supuesto, por ejemplo, que la atmósfera moderna era demasiado ligera para estas criaturas y que, por consiguiente, en el cretáceo la atmósfera debió de haber sido el doble de densa, haciendo posible que el Pteranodon literalmente flotara en el aire[33]. El Pteranodon en realidad no necesitaba esta ayuda atmosférica. En vida el animal pesaba unos 18 kg (algunas estimaciones son de 9 kg), un 25% aproximadamente del peso de un hombre. El Pteranodon había perdido sus dientes, cola y parte de la musculatura de vuelo y sus patas traseras se habían vuelto largas y delgadas. Fue, sin embargo, en los huesos donde se logró la mayor reducción de peso. Los huesos de las alas, columna vertebral y extremidades traseras eran tubulares, como los puntales de soporte de los aviones, que proporcionan resistencia pero disminuyen el peso. En el Pteranodon estos huesos, de hasta 2,5 centímetros de diámetro, no eran otra cosa que espacios aéreos cilindricos limitados por una envoltura ósea externa no más gruesa que una carta. Barnum Brown, del Museo Americano, informó sobre un fragmento del hueso de un brazo de una especie desconocida de pterosaurio del Cretáceo Superior de Texas, en la cual se había alcanzado «la culminación del pterosaurio… la cima de la construcción ligera[34]». ¡Aquí, la tendencia había llegado hasta tal punto que la pared ósea del cilindro tenía el increíble grosor de 0,50 centímetros! En el interior del tubo había puntales óseos al través, no más gruesos que alfileres, que ayudaban a reforzar la estructura; otra «innovación» en el diseño de aviones anticipado por los pterosaurios del mesozoico.
La combinación de un gran tamaño y peso insignificante necesariamente tuvo que provocar cierta fragilidad. Es fácil imaginarse que estos huesos tubulares, delgados como una hoja de papel, que sostenían alas gigantes, posiblemente hicieron difícil el aterrizaje. ¿Cómo podían estas criaturas aterrizar sin romperse todos los huesos? En primer lugar, ¿cómo podían elevarse en los aires? Evidentemente no eran capaces de batir estas alas de 3,6 metros de longitud ensartadas entre tubos delgados como pajitas. Las aves más grandes tienen que alcanzar una cierta velocidad corriendo y batiendo las alas para poder elevarse en el aire, y otras tienen que producir una velocidad de aleteo que casi les permita permanecer flotando antes de poder elevarse en el aire. Para poder permanecer flotando en el aire con una envergadura de alas de 7 metros, el Pteranodon habría necesitado 99 kg de músculos de vuelo tan eficientes como los de los colibríes. Sin embargo, el Pteranodon había reducido su musculatura a unos 3,6 kg, de forma que es inconcebible que pudiera elevarse activamente en el aire[35].
Por lo tanto, el Pteranodon no era una criatura que batía las alas ya que no disponía ni de los músculos ni de la resistencia a la fuerza resultante. Sus alas largas, delgadas como las de los albatros lo revelan como planeador, el planeador más avanzado jamás producido por el reino animal. Con un peso de sólo 18 kg, la carga de las alas era de sólo 0,45 kg por 929 cm2. Esto le proporcionaba una velocidad de descenso incluso inferior a la de los planeadores fabricados por el hombre, cuyas alas deben soportar un peso de, al menos, 1,8 kg por 929 cm2[36]. La relación entre el área de las alas y el peso total en el Pteranodon sólo es superada en algunos insectos. El Pteranodon estaba construido como un planeador con el esternón, articulación del hombro y columna vertebral soldados formando un fuselaje rígido como una caja, capaz de absorber la tensión de las gigantescas alas. El bajo peso, combinado con una enorme envergadura de alas, significaban que el Pteranodon podía planear a velocidades extremadamente bajas sin que hubiera peligro de que se parase. Cherrie Bramwell, de la Universidad de Reading, ha calculado que podía permanecer en el aire con una velocidad de sólo 24 km por hora. De este modo, le habría resultado relativamente fácil elevarse en el aire. Lo único que necesitaba el Pteranodon era una brisa de 24 km por hora para que se pusiera cara al viento, extendiera sus alas y se elevara como un trozo de papel. No era necesario ningún esfuerzo. Si la criatura se hubiese visto obligada a aterrizar sobre el mar, sólo habría tenido que extender sus alas para atrapar el aire y así emerger suavemente del agua.
Parece extraño que un animal, que se había extremado tanto en reducir su peso a un mínimo, desarrollara una cresta ósea, alargada en su cráneo. El hecho de que la cresta estuviera ausente en otros pterosaurios sugiere que no era indispensable para el animal. Se ha sugerido que la cresta era principalmente ornamental[37] pero sencillamente es absurdo imaginarse un ornamento tan engorroso en una criatura voladora tan especializada. Un comentario editorial en el The Aeronautical Journal de 1914, escrito por un científico que no era biólogo, pero que evidentemente estaba familiarizado con la acción de la evolución y economía del diseño de los aeroplanos, puso en ridículo esta idea. «Es obvio que un animal volador que ha reducido su peso al máximo, difícilmente habría desarrollado una larga cresta a costa de, o sin efecto sobre, su capacidad para volar. Es decir, la cresta debió de tener alguna ventaja aerodinámica para su propietario para que pudiera evolucionar en la medida que lo hizo. No es difícil ver que este tipo de cresta pudo ser muy útil desde el punto de vista de la estabilidad.
A todos aquellos que estén familiarizados con la extrema importancia que posee la disposición de las aletas para la estabilidad de los aeroplanos, no será necesario explicarles que, cuando el Pteranodon dirigía su pico hacia abajo, movía una gran área de aleta (el mismo pico) hacia atrás y abajo[38]». Sin embargo, cuando el pico bajaba, concomitantemente se elevaba la cresta para compensar el efecto: el Pteranodon estaba equipado con un mecanismo automático de contracción. Más tarde, se demostró que la cresta, en realidad, ahorraba peso. Los músculos adicionales del cuello necesarios para mantener el pico recto cara al viento, habrían excedido en mucho el peso de la cresta[39]. Esto ha sido demostrado mediante experimentos con tanques de descarga y túneles de viento con modelos a escala reducida de cabezas de Pteranodon con cresta y sin cresta. Estos experimentos también revelan otra función de la cresta. Cuando se eliminaba la cresta, la cabeza era desviada constantemente hacia un lado por la fuerza de la corriente y, en vida, los músculos del cuello habrían tenido que actuar continuamente para mantener la cabeza dirigida hacia delante. Cuando se volvía a colocar la cresta en su sitio, se producía una corrección automática si la cabeza se desviaba oblicuamente a la dirección del flujo de aire. En otras palabras, el pico y la cresta actuaban conjuntamente como una veleta estando dirigidos siempre en dirección al viento. La cresta probablemente también funcionaba como un timón en un planeador para evitar movimientos de derrape laterales durante los giros[40].
El vuelo de los pterosaurios siempre ha llamado la atención de los ingenieros aeronáuticos. En 1914, el año de la muerte de Wilbur Wright, se publicó un informe de un equipo de trabajo muy curioso compuesto por D. M. S. Watson, el especialista en fósiles de reptiles del University College de Londres y E. H. Hankin, un experto en aeronáutica. En éste analizaban la complicada serie de articulaciones del hombro, codo, muñecas y nudillos en el Ornithodesmus del cretáceo inferior, con una envergadura de 4,5 metros. En vista de la delicada naturaleza de los huesos, y de la ausencia de extensos músculos de vuelo en estos pterosaurios gigantes, Watson y Hankin llegaron a la conclusión de que «no volaban peor que otras aves sino que volaban de forma más científica[41]». La complicada serie de articulaciones les proporcionaba un incremento de la maniobrabilidad y, posiblemente, les permitía frenar suavemente mientras se elevaban hacia el cielo.
Recientemente, se ha atribuido una función similar al enigmático hueso pteroideo, el pequeño sobrehueso en el dedo del ala para el cual no se había encontrado ninguna función. Cherrie Bramwell sugiere que sostenía una membrana que se extendía hasta el cuello. Cuando la criatura se eleva en el aire, el hueso podía ser levantado y la membrana actuaba como un elevador, como en los aviones modernos. Cuando el animal quería posarse sobre tierra, el hueso era descendido para reducir la velocidad a un mínimo sin crear turbulencias y paradas del vuelo. El Pteranodon podría así aterrizar sin peligro de lesionarse sus frágiles huesos.
Cuando Cherrie Bramwell introdujo los datos del Pteranodon en el ordenador de la Escuela de Aeronáutica, se descubrió hasta qué punto estos planeadores naturales estaban avanzados. El ordenador estaba programado para examinar el comportamiento de vuelo de los planeadores construidos por el hombre y no necesitó ningún ajuste para el Pteranodon[42]. Parece ser que con velocidades superiores a 18 nudos (aproximadamente 32 km por hora), el Pteranodon podía girar sus alas hacia atrás en forma de V (a diferencia de los murciélagos podía hacerlo debido a que sus alas estaban sostenidas por una larga serie de huesos que podían ser flexionados en los nudillos). Esto provocaba una disminución de la velocidad de descenso y es el principio empleado por los aviones a reacción con alas de geometría variable: un principio descubierto de nuevo, después de haber estado dormido durante 70 millones de años.
Los sedimentos marinos que contienen esqueletos de Pteranodon también albergan una gran diversidad de reptiles marinos, especialmente plesiosaurios y mosasaurios. Algunos de los depósitos que contenían pterosaurios se formaron a cientos de kilómetros de distancia de la costa del mar más cercana, que invadió Tejas y Kansas en la era cretácea. Esto sugiere que el Pteranodon podía recorrer grandes distancias lejos de tierra como los albatros[43]. Evidentemente, tuvo una existencia idílica en el aire dependiendo sólo de los peces que recogía. Al igual que los pelícanos modernos, poseía una bolsa en el cuello (que en ocasiones puede ser observada en los fósiles con luz ultravioleta), en la cual guardaba los peces. Un espécimen en el Museo Americano tiene los esqueletos de dos especies de peces que aún estaban en su bolsa, dando fe de sus gustos dietéticos.
El modo cómo recogía los peces nos plantea algunos problemas. Existen algunos murciélagos que también se alimentan de peces. Al igual que los Pterosaurios, poseen enormes patas traseras con garras que cuelgan cuando están volando. El murciélago alano (Noctilio leporinus), de cuyos hábitos conocemos algo, vuela por encima del agua al atardecer y desciende para coger los peces con sus garras. Éste probablemente también es el método empleado por los otros murciélagos pescadores aunque sus costumbres son poco conocidas. Existe un murciélago pescador en California (Pizonyx vivesi), que durante el día vive junto a petreles en grietas rocosas, del cual, ya antes de haber sido observado durante la noche, se sospechaba que era un pescador debido a sus largas garras traseras. Esto fue confirmado posteriormente por un análisis del contenido de su estómago que reveló que sólo contenía peces. Sin embargo, ¿era posible que el Pteranodon dejara colgar sus garras en el mar para coger peces? Si éste era el método, y vivía a cientos de kilómetros de tierra, ¿cómo llegaban los peces hasta su boca? Es bastante más probable que el Pteranodon planeara por encima de las crestas de las olas atrapando los peces que vivían en la superficie del agua con su pico extremadamente largo, montado a un cuello móvil. Quizá se posaba sobre la superficie para coger los peces. Últimamente, se ha sugerido que el dedo del ala podía girar produciendo con el ala un plano en forma de M que iniciaría la inmersión inmediata[44]. Si esto fuera posible, debió de haber sido desde una altura bastante baja a fin de reducir el impacto de los extremadamente ligeros huesos con el agua. Una generación anterior, quizá más influenciada por los aeroplanos como instrumentos de destrucción, vio el Pteranodon desde otro punto de vista. Barnum Brown se refería a los pterosaurios como «los aeroplanos de la época prehistórica». En un artículo publicado en octubre de 1943, Brown veía un paralelismo entre las aterradoras técnicas aéreas contemporáneas y el estilo de vida de los primitivos pterosaurios. El «bombardero de inmersión». Pteranodon, como lo llamaba Brown, era la máquina voladora más desarrollada de su época. La cresta era «una especie de timón que le era útil al animal cuando, con las alas dobladas, cruzaba el aire como un rayo en busca de presas… El cuello era moderadamente largo, fuerte y flexible. Presentaba una notable serie de articulaciones adicionales nunca vistas en los huesos del cuello de otros animales, que proporcionaban una gran fuerza al pico para asestar golpes[45]». Sin embargo, Barnum Brown había sido influido excesivamente por los sucesos siniestros de su época. El incremento de la sofisticación de la guerra aérea le había llevado a investir al Pteranodon de una agresividad que en vida no existía. Es poco probable que el frágil Pteranodon pudiera haber resistido el choque de precipitarse al agua desde cualquier altura. A diferencia de las gaviotas, que pueden recoger sus alas, el Pteranodon no tenía cóndilos y carillas articulares en la articulación del hombro, de forma que no podía recoger sus enormes alas y, al estar éstas extendidas, no podrían haber resistido el impacto. ¡Tal como sugirió el mismo Watson, en 1914, en una época en la cual la guerra era un poco menos sofisticada, y cuando los aviadores sabían lo que significaba la fragilidad en el aire, probablemente el enorme pterosaurio se habría desintegrado al intentar enderezarse después de una inmersión[46]! En lugar de ser el diabólico bombardero buceador de Barnum Brown, el Pteranodon era una criatura grácil, muy inteligente, que se dedicaba a volar plácidamente sobre los grandes mares del cretáceo.
El Pteranodon se pasaba los días planeando sobre el océano en busca de peces que vivían en la superficie. Tradicionalmente, se pensaba que estos Pterosaurios gigantes eran negros con alas curtidas como las de los murciélagos. Desde que sabemos que el «demonio peludo» de Sharov, Sordes pilosus, tenía un grueso abrigo de piel, y que las membranas de sus alas también estaban cubiertas de pelo parece más probable —aunque a primera vista resulte sorprendente— que el Pteranodon estaba cubierto de pelo blanco. Esta piel le habría proporcionado a la criatura un parecido aún mayor con el albatros. Las gaviotas son blancas, lo cual constituye una forma de camuflaje; al presentar un perfil menos evidente para los peces, evitan ahuyentar al potencial alimento. Esto habría constituido un problema aún mayor para el pescador Pteranodon. Sus enormes alas habrían tapado el sol a medida que sobrevolara el agua y unas alas negras habrían agravado seriamente la situación.
Decir que, como criatura voladora, el Pteranodon era grande sería una descripción insuficiente. De hecho, existen razones fundadas para creer que se trataba del animal más grande que jamás pudo elevarse en los aires. Con cada incremento del tamaño y, por lo tanto, también del peso, un animal volador necesita un incremento concomitante de fuerza (para batir las alas en los animales aleteadores y para sostenerlas y manipularlas en los planeadores). Sin embargo, la fuerza es suministrada por músculos que a su vez añaden aún más peso a la estructura. Cuanto más grande se hace un volador, más desproporcionadamente pesado se vuelve por la adición de su propio suministro de fuerza. Llegará un punto en el cual el peso será demasiado grande para permitir que la máquina se eleve en el aire. Los cálculos realizados, en relación con el tamaño y la fuerza, sugieren que el peso máximo que puede alcanzar un vertebrado volador es de aproximadamente 22 kg: por esta razón, se piensa que el Pteranodon, y su homólogo jordano Titanopteryx, ligeramente más grande pero menos conocido, eran los animales voladores más grandes[47].
Sin embargo, en 1972 se produjo el primero de una espectacular serie de hallazgos que sugieren que debemos cambiar drásticamente nuestras ideas acerca del tamaño máximo permisible en vertebrados voladores. A pesar de que las excavaciones aún prosiguen, los descubrimientos hechos durante tres temporadas —desde 1972 hasta 1974—, por Douglas A. Lawson de la Universidad de California, han revelado esqueletos parciales de tres pterosaurios extremadamente grandes en el Parque Nacional de Big Bend, en el condado de Brewster, Texas. Estos esqueletos pertenecen a criaturas que hacen que incluso el Pteranodon parezca pequeño. Lawson halló los restos de cuatro alas, un largo cuello, extremidades posteriores y maxilares carentes de dientes en yacimientos no marinos; se cree que estos antiguos sedimentos sepultados se originaron debido a aluviones en llanuras sujetas a inundaciones. El inmenso tamaño de los pterosaurios de Big Bend, que en el mundo paleontológico se conocen como «los 747» o «Jumbos», podrían ser calibrados colocando los húmeros de Texas al lado de los de un Pteranodon: el húmero del «Jumbo» tiene el doble de longitud que el del Pteranodon. ¡El ordenador de Lawson estimó una envergadura de más de 75 metros para este planeador! «No es ninguna sorpresa —dijo Lawson al anunciar el animal en Science en 1975— que los restos de esta criatura fueran hallados en Texas[48]».
A diferencia del Pteranodon, estas criaturas fueron encontradas en rocas que se formaron a 400 kilómetros en el interior de la costa cretácea. La ausencia de yacimientos de lagos en los alrededores, habla en contra de que estos pterosaurios hubieran sido pescadores. Lawson sugiere que habían sido carroñeros, hartándose de montones de carne podrida abandonados después del desmembramiento del cadáver de un dinosaurio. Quizás, al igual que los buitres y los cóndores, estos Pterosaurios volaban por encima del cadáver esperando su turno. Después de haberse posado sobre el cadáver, sus picos sin dientes sólo les habrían permitido alimentarse de los blandos órganos internos. Cómo pudieron elevarse en el aire después de haberse hartado de comida es un enigma. El animal no podría haber batido alas de un tamaño tan extraordinario estando en el suelo. Debido a que los pterosaurios eran incapaces de correr a fin de emprender el vuelo, debieron de elevarse verticalmente en el aire. Las palomas sólo pueden despegar verticalmente, reclinando sus cuerpos y batiendo las alas delante de ellos. En el caso de que fueran aleteadores, ¡los pterosaurios de Texas habrían necesitado tener patas muy largas en forma de zancos a fin de elevar el cuerpo lo suficiente como para permitir que sus alas de 7 metros de longitud despejaran el suelo! La principal objeción, sin embargo, sigue siendo la ausencia de una musculatura adecuada para una operación de este tipo. ¿Es que la única solución es suponer que estas criaturas, con las alas completamente extendidas y los elevadores alzados, fueran levantadas pasivamente del suelo por el viento? Si Lawson tuviera razón, y los pterosaurios de Texas fueran carroñeros (lo cual comienzo a dudar), se podría prever otro problema. La existencia de cadáveres de dinosaurios implica la presencia de dinosaurios. Los gigantescos y torpes pterosaurios eran vulnerables a ser atacados estando en el suelo. Así que, ¿cómo podían escapar de los formidables dinosaurios? Estando a merced de las corrientes de viento, el despegue habría sido una empresa arriesgada[49]
Los exóticos pterosaurios de Lawson plantean algunas cuestiones intrigantes. Sólo la prosecución de la investigación nos dará las respuestas.