III. LA CARRERA ES PARA LOS VELOCES, LA LUCHA PARA LOS FUERTES
Esta, aún muy controvertida, idea inmediatamente comenzó a tener partidarios que veían en ella uno de los conceptos más fructíferos y productivos. Sugería hacer nuevas preguntas y plantear nuevas formas de abordar la cuestión que, inesperadamente, mostraban al dinosaurio bajo un punto de vista completamente nuevo.
Por una notable coincidencia, este nuevo enfoque había surgido simultáneamente al otro lado del Atlántico. Armand de Ricqlés en la Universidad de París había llegado independientemente a la misma conclusión de Ostrom en unos pocos meses aunque su modo de abordar el problema había sido bastante distinta. (Este tipo de feliz coincidencia es una característica recurrente de los descubrimientos científicos. En ciertas épocas, parece que las ideas nuevas floten en el aire esperando que alguien tenga el valor de cogerlas; recuérdese la lucha entre Newton y Leibniz acerca de quién inventó el cálculo, o el sobresalto de Darwin cuando recibió un resumen de Wallace acerca de su propia teoría de la evolución mediante selección natural). Así, en 1969, después de haber estudiado la estructura interna de muchos tipos de huesos fósiles y recientes, Ricqlés, sugirió que fisiológicamente los dinosaurios eran más parecidos a los mamíferos que a los lagartos. El hueso, aparte de su obvio papel como andamio interno, es una estructura activa, en crecimiento y fisiológicamente importante: en la médula se fabrican células sanguíneas y el tejido óseo actúa como un importante depósito de calcio (las hormonas desencadenan la liberación del calcio del hueso y lo envían a los lugares de la contracción muscular donde desempeña un importante papel). Los animales activos con un alto nivel metabólico y sistemas fisiológicos acelerados exigirán un mayor rendimiento de los huesos, lo cual se reflejará en su fina estructura. Ricqlés se dio cuenta de que la estructura ósea era un sensible indicador del nivel de actividad de un animal y que, en los huesos fósiles, disponíamos de todos los ingredientes para evocar la fisiología de los animales muertos hacía mucho tiempo. En los mamíferos y las aves, los vasos sanguíneos que penetran en los huesos largos son numerosos y están densamente agrupados (permitiendo un mayor transporte de materiales hacia y desde regiones activas); en los reptiles, los vasos sanguíneos son escasos. Los conductos de Havers, los lugares que controlan el intercambio de calcio entre el esqueleto y la sangre, son menos numerosos en los huesos de los reptiles que en los huesos de los mamíferos, y su falta limita la velocidad de intercambio. Ricqlés afirmó que, los huesos de los dinosaurios se parecen a los huesos de los mamíferos, pero que se diferenciaban claramente de los huesos de reptiles y anfibios de sangre fría. Interpretó esta sorprendente «convergencia» como prueba de la existencia de un metabolismo activo, y que los dinosaurios posiblemente eran animales de sangre caliente[1].
Estos estudios independientes de Ostrom y Ricqlés convirtieron a los mamíferos en el nuevo paradigma para los dinosaurios, eliminando así las limitaciones asociadas al obsoleto modelo de lagarto. Estos nuevos modelos centraron la atención sobre aspectos hasta entonces inexplorados de los dinosaurios, aspectos que nunca hubiesen salido a la luz bajo el antiguo orden. La tarea de estudiar a fondo todas estas nuevas ideas estuvo en manos de un estudiante de Ostrom en Yale, Robert T. Bakker. Bakker se llevó consigo estas radicales ideas al Museo de Zoología Comparada en la Universidad de Harvard y fue aquí, en el museo de Louis Agassiz, donde analizó las implicaciones de la endotermia para los dinosaurios en una serie de artículos escritos a principios de la década de 1970. Esta nueva forma de abordar el problema también provocó que los problemas existentes fueran analizados desde una nueva perspectiva. Frecuentemente estos problemas habían pasado inadvertidos y las paradojas habían sido aceptadas sin levantar crítica alguna. Las desventajas fisiológicas de los lagartos no podían ser ignoradas, y aunque se reconocía que los reptiles vivos están impedidos por su limitada capacidad para correr o incluso para mantener una posición erguida, se suponía que los dinosaurios realizaban prodigiosas proezas de gran fuerza y resistencia. Aun a pesar de que estas limitaciones se habrían incrementado desproporcionalmente en un lagarto del tamaño de un elefante, se continuaba pensando que estos formidables carnívoros se habían pasado la vida persiguiendo activamente a sus presas, mientras que, a su vez, los herbívoros habían sido capaces de escapar de sus garras. Si un pequeño lagarto, que pesa pocos gramos, solamente es capaz de tener breves períodos de actividad, ¿cómo un carnívoro de ocho toneladas de peso como el Tyrannosaurus habría podido aguantar una lucha feroz y prolongada con su presa? La muerte de un Triceratops en las garras del tiranosaurio no podía haber sido tan instantánea como lo es la de una mosca atrapada por un lagarto. Un dinosaurio grande tenía un volumen aproximadamente un millón de veces superior al de un lagarto pequeño, y aún así se creía que éste había cazado sus presas y mantenido combates a muerte. ¿De dónde sacaban la energía?
Una forma de salir de esta paradoja era suponer, como hicieron algunos científicos en el caso de los gigantes carnívoros, que eran demasiado grandes y pesados como para haber cazado activamente sus presas y luego haber luchado. Lawrence Lambe, el descubridor del monstruo canadiense Gorgosaurus, era el más ferviente partidario de esta teoría. El descubrimiento de un nuevo dinosaurio carnívoro siempre es un acontecimiento. Los carnívoros no eran ni con mucho tan abundantes como sus inofensivos contemporáneos herbívoros en la época mesozoica, pero al ser bestias tan imponentes, su escaso número pronto se veía compensado por el interés que suscitaba en sus descubridores. Frecuentemente, una comunidad de una determinada localidad tendrá una serie de herbívoros, pero sólo a un único carnívoro gigante para tenerlos a raya. Al explorar la región canadiense del río del Ciervo Rojo de Alberta, a principios del siglo, se descubrieron muchos herbívoros nuevos, y en 1913 Lambe entró en posesión de un carnívoro completamente nuevo. El Gorgosaurus era un gigante del cretáceo tardío: alcanzaba una longitud de 8,8 metros y una altura de 3,3 metros. Aunque estaba armado con los obligatorios dientes y maxilares fuertes, el gorgosaurio tenía una constitución más ligera que el Tyrannosaurus, con patas traseras más delgadas y más gráciles. Con todo, obviamente pesaba algunas toneladas y Lambe pensaba que la criatura había pasado la mayor parte de su vida tumbada en el suelo. Podía caminar erguida si la situación lo requería, y en este caso la cola actuaba como contrapeso al tronco y el peso del cuerpo era soportado por las patas traseras. Sin embargo, esto sucedía muy raras veces, provocado sólo por un motivo muy fuerte. En posición postrada, sentada o agachada, el peso del gorgosaurio pasaba directamente al suelo a través del cinturón pélvico. «Esta posición de reposo —aventuró Lambe—, y particularmente la postura con el cuerpo acostado en toda su longitud (cuando la criatura estaba completamente estirada con la cabeza apoyada sobre el suelo) probablemente eran las más frecuentemente adoptadas por un reptil con la forma, y naturaleza supuestamente perezosa del gorgosaurio». Extrañamente, los largos dientes aserrados en los maxilares del gorgosaurio estaban completamente nuevos sin estrenar, a pesar de que el ejemplar no era joven. Las puntas de los colmillos eran casi perfectas, con delicados bordes dentados en los agudos filos cortantes. Lambe dedujo que los alimentos ingeridos por este carnívoro eran blandos y no abrasivos, y que la presa no podía haber ofrecido resistencia alguna.
Por consiguiente, se cree que el gorgosaurio se limitaba a alimentarse de los cadáveres de animales que no habían sido matados recientemente, que no era un intrépido cazador sino un animal que se alimentaba de carroña que desempeñaba su papel en la Naturaleza y sin duda sus servicios eran requeridos a menudo, si tenemos en cuenta la inmensa cantidad de tracodontes, ceraptosianos, estegosaurios y otros dinosaurios y reptiles que vivían y morían en este período particular del cretáceo[2].
Sumergido, como Lambe creía, en montones de potencial alimento que se desplazaban lentamente, el Gorgosaurus no necesitaba ser un depredador activo. A su alrededor debían de haber muerto de forma natural todo tipo de herbívoros. La bestia engullía «los cadáveres que encontraba o con los que se tropezaba en sus excursiones provocadas por el hambre». El monstruoso gorgosaurio era un animal de carroña casi inofensivo. Arrancaba los blandos tejidos del cadáver con sus garras antes de agacharse sobre la carne en proceso de descomposición para consumir los sabrosos tejidos. Al menos, eso es lo que pensaba Lambe.
Desgraciadamente para la teoría de Lambe, los dientes se pierden o desgastan, no durante el acto de matar, sino al arrancar la carne de los huesos. El Museo Americano tiene el esqueleto completo de un brontosaurio, cuyos huesos de la cola han sido roídos por un alosaurio. Al comparar un maxilar de alosaurio con las muescas de los dientes se vio que éstas encajaban exactamente. Obviamente, un Allosaurus había tenido un banquete con el cadáver del brontosaurio. Es más, durante este proceso había perdido varios dientes que fueron encontrados al lado de la cola del brontosaurio cuando fue extraída de las rocas[3]. Resulta incomprensible por qué los dientes del gorgosaurio canadiense están como nuevos, ya que por el mero hecho de comerse el cadáver se hubiesen desgastado. Naturalmente, esto no puede ser utilizado como prueba para demostrar un comportamiento cobarde del gorgosaurio. La serie de armamentos que llevaba era tan temible como la de un alosaurio o tiranosaurio. La alimentación a base de carroña parece haber sido una ocupación poco probable para esta bestia; tiene el aspecto como si estuviese hecho para la agresión activa. Además, la mayoría de los investigadores que estudiaban a los grandes carnívoros como el Allosaurus y Tyrannosaurus de Norteamérica y el Tarbosaurus, el homólogo mongol del tiranosaurio, no compartían el escepticismo de Lambe, y en lugar de ello preferían creer en la evidencia de las garras y maxilares, éstas indicaban que se trataba de formidables asesinos. En la vida (y la muerte), naturalmente las cosas nunca están tan claras: un león puede alimentarse de carroña y una hiena en ocasiones matar. Sería poco realista imaginarse a un gorgosaurio o tiranosaurio renunciar a una comida gratuita si se encontraran con un animal muerto. No obstante, parece ser más probable que, al igual que los leones, se tratara principalmente de depredadores activos.
El Tyrannosaurus rex, el «rey de los lagartos tiranos», era la expresión extrema de su tipo. Su impresionante tamaño, de poco menos de 12 metros de longitud si aceptamos la estimación revisada de Newman, y 6 metros de altura cuando el animal estaba erguido sobre sus patas traseras, no deja lugar a dudas de que se trataba del rey mesozoico de las bestias. Fue encontrado por primera vez como resultado de un intenso período de actividad en el Museo Americano a finales del siglo pasado. Bajo la presidencia de Henry Fairfield Osborn, una gran cantidad de expediciones bien equipadas fueron enviadas a las Montañas Rocosas, que como resultado hicieron descubrimientos tan prolíficos que aún ahora estamos cosechando los beneficios. El Tyrannosaurus surgió de su tumba de Hell Creek en Montana, en 1902, cuando el equipo de Barnum Brown estaba explorando la región en busca de dinosaurios. Brown, acompañado de Lull, localizó el cráneo, maxilares, columna vertebral y patas traseras de la bestia, pero la dura piedra caliza circundante del cretáceo superior obligó a los equipos del museo a volver durante dos o tres años consecutivos para desenterrar por completo al animal mientras que Lull, ya de vuelta en el museo, preparaba el esqueleto para montarlo. El Tyrannosaurus sorprendió a todo el mundo. El Allosaurus, a pesar de su gran tamaño, parecía pequeño al lado del más grande de todos los tiranos. Un colega de Osbom en el museo, William Diller Matthew, intentó hacer un dibujo preliminar de la bestia y para destacar sus dimensiones de la forma más dramática, la colocó junto a un diminuto esqueleto humano. Este espécimen, y gran parte de otros dos esqueletos hallados en las sucesivas incursiones a Hell Creek, fueron estudiados por Osborn, quien, en reconocimiento a esta criatura que había sido el carnívoro terrestre más grande que jamás pisó la Tierra, la bautizó con el nombre de Tyrannosaurus rex[4]. Una criatura, además, que estaba construida sólidamente e indudablemente para luchar, con largos dientes letales y unos terroríficos maxilares y unas garras de 20 centímetros de longitud que armaban los dedos de los pies. Al igual que Cope, que al hallar al Laelaps carnívoro en la misma área que al hadrosaurio, se había visto obligado a considerar a ambos como enemigos, Osbom sugirió que el tiranosaurio había sido el enemigo implacable del Triceratops cornudo. La armadura del herbívoro tricornio había alcanzado tal grosor que el Tyrannosaurus había desarrollado dientes de 15 centímetros de longitud para penetrar en ella y unos maxilares de 1,20 metros de longitud para poder engullir los pedazos de carne.
El estudio del cráneo del Tyrannosaurus revela sus hábitos depredadores. Debido a que las patas delanteras no servían para desgarrar la carne de la forcejeante presa, el cráneo había asumido la función principal de agresión. Consiguientemente, había crecido desproporcionalmente en comparación con el tamaño del animal y estaba armado con una serie de dientes curvados, aserrados, parecidos a lanzas paleolíticas. La longitud de sus maxilares permitía una mordedura realmente efectiva y el Tyrannosaurus era así capaz de engullir considerables masas de carne de dinosaurio. El Allosaurus, más pequeño incluso, superaba al tiranosaurio en su capacidad de tragar cantidades considerables de presas. En la región de la sien, el hueso cuadrado que se articulaba con el maxilar inferior era ligeramente móvil mientras que los dos maxilares y el techo del cráneo podían flexionarse en sus puntos centrales. Todo ello constituía una serie de adaptaciones notables que hacían que la boca se pudiera abrir más y el Allosaurus pudiera engullir vorazmente enormes masas de carne.
Dos de los tiranosaurios rescatados por Barnum Brown en Montana, fueron restaurados en la sala cretácea del Museo Americano en Nueva York. Las posturas adoptadas por Osborn para sus reconstrucciones eran las dibujadas por el director del departamento de reptiles del parque zoológico de Nueva York, tras un intensivo estudio de los movimientos de los lagartos. Estas restauraciones, dijo Osbom, representaban «animales antes del salto convulsivo y mordedura que distingue la lucha de los reptiles[5]».
Barnum Brown describió la escena más vivamente.
Son las primeras horas de la mañana en las orillas de un lago del cretáceo hace tres millones de años. (En la época de Barnum Brown —este artículo fue escrito en 1915— se le concedía un período mucho más corto a la duración de la historia de la Tierra: actualmente se cree que desde el final del cretáceo han transcurrido 64 millones de años). Un dinosaurio herbívoro, un Trachodon, que se ha aventurado fuera del agua para conseguir un desayuno a base de vegetación suculenta, ha sido apresado y devorado parcialmente por un Tyrannosaurus gigante, carnívoro. Mientras este monstruo se agacha sobre el cadáver, descuartizándolo atareadamente, otro Tyrannosaurus es atraído a la escena. Se aproxima irguiéndose para agarrar al más afortunado cazador y disputarle la presa. La figura agachada, de mala gana deja de comer y acepta el desafío levantándose parcialmente para saltar sobre su adversario.
El momento psicológico de esta tensa inercia, antes del combate, fue escogido para mostrar mejor las posiciones de los miembros y de los cuerpos y para describir un incidente en la historia de la vida de estos reptiles gigantes[6].
Cope había sentado el precedente colocando a sus Laelaps agarrados uno a la garganta del otro, de modo que Brown le siguió el ejemplo enfrentando a un tiranosaurio con otro. Sin embargo, esta agresión intraespecífica parece improbable. El hecho de que muchos autores hayan asumido que las limitadas capacidades mentales de los dinosaurios depredadores hacían que éstos atacaran todo lo que se moviera, es razón de más para creer que existía algún tipo de comportamiento territorial entre los carnívoros más grandes, similar al que actualmente existe entre los leones. Los dinosaurios depredadores más pequeños probablemente cazaban en manadas como los lobos[7], existiendo en este caso muy probablemente algún tipo de jerarquía —una jerarquía social de los dinosaurios— para evitar que se produjeran estas reyertas destructivas que Cope y Brown habían imaginado.
Brown pensaba que el Tirannosaurus era «una criatura poderosa, activa y de movimientos ágiles si la ocasión lo requería». De hecho, cuando Brown desenterró un esqueleto de Gorgosaurus en la región del río del Ciervo Rojo en Alberta, una década más tarde, lo envió al museo para que fuese restaurado en una postura como si estuviese corriendo, como cazando una manada de dinosaurios de pico de pato[8].
Osborn y Matthew, conscientes de la limitada capacidad del lagarto para mantener una agresión activa, eran más reservados en sus afirmaciones concernientes a las agresivas luchas y agilidad de estos carnívoros de gran tonelaje. Los lagartos matan con rapidez, una embestida y el destino del insecto está sellado, y Matthew se imaginaba algo parecido en el antiguo mundo. «En lo referente a las probables costumbres —dijo del Allosaurus (cuyo nombre significa «reptil saltador», un legado de la época de Cope)—, es sensato deducir que era un depredador, activo y poderoso, adaptado a la vida terrestre. Sus métodos de ataque y lucha deben de haber sido más parecidos a los de los reptiles modernos que a los de los mamíferos carnívoros más inteligentes». Se imaginaba al Allosaurus echado en el suelo a la espera (¿ciertamente uno de los métodos inteligentes de los depredadores sofisticados actuales?), «luego un repentino y rápido ataque, una feroz mordedura de los enormes maxilares y un salvaje ataque con dientes y garras hasta que la víctima había sido descuartizada o engullida entera[9]». Era como si un carnívoro de ocho toneladas de peso pudiera imitar a un lagarto cuando ataca su presa y se retira rápidamente para volver a recuperar sus fuerzas. Esto, naturalmente, es muy improbable. El enorme volumen del Allosaurus y de sus enemigos habrían impedido una victoria de este tipo. Las presas no eran diminutas criaturas inofensivas, como lo son las presas del lagarto, ya que en el caso del Tirannosaurus cabe suponer que se trataba de un ceratopsiano armado tricornio o de un hadrosaurio de pico de pato más ágil.
Sorprendentemente, Matthew se contradijo a sí mismo cuando volvió a revisar las probables costumbres del tiranosaurio. «Su volumen imposibilita la rapidez y agilidad —declaró, sin advertir que esta afirmación también debía haber sido aplicable al Allosaurus que a su vez alcanzaba los 10 metros de longitud—. Debe de haber estado diseñado para atacar y devorar a los pesados y lentos dinosaurios con cuernos y armadura con los que han sido encontrados sus restos, y cuyas macizas corazas y armas de defensa se emparejaban bien con sus dientes y garras. El momento implicaba una acción aparentemente lenta y pesada de su enorme cuerpo, una inercia de movimientos, difícil de iniciar y difícil de cambiar o parar. Estos movimientos se diferencian mucho de la ágil rapidez que asociamos con las bestias depredadoras». «Ningún animal de las dimensiones de un tiranosaurio podía saltar sobre su presa —concluyó—, y un avance rápido hacia la lucha bien podría haber acabado en un inevitable empalamiento en los puntiagudos cuernos del Triceratops». Entonces, ¿cómo describía Matthew los «combates de los titanes del antiguo mundo»? Probablemente eran luchas largas contra adversarios bien armados, luchas que dejaban poco respiro a los protagonistas para recuperar sus fuerzas. ¿Podría un reptil de sangre fría, fisiológicamente ineficiente, de ocho toneladas de peso haber resistido estas luchas con un metabolismo que producía poca energía? Un tiranosaurio ectotérmico, de hecho, no habría tenido que retirarse antes que un lagarto, cuyo diminuto peso, después de todo, somete los músculos a un esfuerzo mucho menor?
W. E. Swinton, al discutir las costumbres de los tiranosaurios en 1934, se imaginaba las luchas de estos primitivos reptiles como enfrentamientos prolongados y feroces. «Los Tyrannosaurus —dijo de estos superdepredadores del cretáceo—, no podían haber sido otra cosa que torpes y desgarbados gigantes que luchaban contra formas de peso similar».
No era una criatura astuta o inteligente sino que en gran parte se guiaba por el instinto y las reacciones automáticas frente a los estímulos visuales y olfativos, si bien la larga práctica a través de incontables ancestros carnívoros había provocado que esta coordinación alcanzara un elevado nivel de eficacia. A pesar de ello, las limitaciones mecánicas del cuerpo se oponían a los repentinos saltos, persecuciones veloces o contiendas entre dos inteligencias que caracterizan a las luchas de los mamíferos; y, sin duda, las contiendas del mundo cretáceo, si las pudiéramos ver ahora, parecerían tener la rigidez de las actividades de aficionados más que la suavidad de los profesionales aunque las disputas eran reales y terribles[10].
La deducción de la función de las estructuras (tales como dientes y garras) a partir de su forma, sólo es uno de los medios, si bien el principal, para demostrar que los dinosaurios terópodos (carnívoros) eran agresores activos. A finales de la década de los años 1930, Roland T. Bird, un explorador que buscaba dinosaurios para el Museo Americano y compañero de Barnum Brown, causó bastante revuelo con sus descubrimientos de dinosaurios. Bien, no exactamente de los mismos dinosaurios sino de sus huellas, dejadas para la posteridad cuando los antiguos leviatanes cruzaron superficies fangosas. En aquellas ocasiones, en las que el fango era lo suficientemente duro quedaron impresas en el suelo huellas bien definidas que se llenaron con sedimentos de aluviones. Cuando el suelo se elevó y las rocas sobrepuestas se erosionaron millones de años más tarde, el esquisto del interior de las huellas se desgastó dejando impresiones casi perfectas de las antiguas huellas. La ciencia que se ocupa de las huellas fósiles, o iconografía, recibió un inmensurable impulso debido a las numerosas huellas encontradas por Bird en los Estados de las Montañas Rocosas.
Hacia finales de noviembre de 1938, Bird llegó al escenario donde se encontraban las enormes huellas de carnívoros con tres dedos —perfectamente impresas en los lechos—, después de haber visto algunas planchas de piedra que contenían huellas fósiles y que estaban a la venta en un tenderete de recuerdos indios. Las huellas de dinosaurios con tres dedos no eran raras, ya que se habían descubierto muchas en el siglo transcurrido desde que Edward Hitchcock coleccionara sus huellas de «aves» en el valle del Connecticut para su museo en el Amherst College. Hitchcock había descubierto las huellas de dinosaurios primitivos que habitaban la región de Nueva Inglaterra en la época triásica, mientras que Bird estaba tratando con piedras mucho más jóvenes, de aproximadamente 120 millones de años de antigüedad en Glen Rose a 128 kilómetros al sudoeste de Fort Worth en Texas. Glen Rose era territorio virgen para las expediciones del Museo Americano de Historia Natural. Bird, viendo una piedra de mampostería que contenía la impresión de una huella de un carnívoro de medio metro de longitud perteneciente al edificio de la administración de justicia local, se dio cuenta que la región sería provechosa. Parece ser que durante mucho tiempo la ciudad había dado por supuesto que estas «huellas eran de hombre». Las huellas eran muy numerosas en el lecho del río Palaxy, unos kilómetros río arriba donde las aguas atraviesan las marismas del cretáceo inferior y dejaban al descubierto las huellas. Bird pasó los últimos días de aquella temporada excavando estas huellas de carnívoro con garras de águila, pero también desenterró, casi de forma accidental, la primera sucesión conocida de huellas con cuatro dedos de brontosaurios gigantes que se movían pesadamente. Estas huellas de saurópodos de 96 centímetros de longitud lo hicieron volver a toda prisa a Glen Rose en la siguiente temporada. Los equipos del Museo Americano y de la Universidad de Texas extrajeron de este lugar unas 40 toneladas de planchas que contenían huellas, proceso durante el cual encontraron una sucesión de huellas particularmente espectaculares. Un imponente saurópodo había atravesado las marismas sin darse cuenta que un carnívoro lo estaba siguiendo de cerca. Cuando el brontosaurio torció hacia la izquierda, el gran carnívoro lo siguió. La tensión iba aumentando incluso entre los trabajadores más impasibles a medida que se iba exponiendo el drama y descubriendo más huellas. Existía incluso cierto grado de amistosa rivalidad al desenterrar las huellas y se hacían especulaciones sobre el resultado de esta cacería prehistórica. ¿Podría escapar el brontosaurio a la seguridad de las profundidades o se encontraría su esqueleto medio comido en algún lugar más adelante? El decepcionante final de la persecución se produjo cuando la roca quedó enterrada debajo de un muro de piedra caliza. El resultado sigue sin conocerse[11]. ¿En primer lugar, hubo realmente una cacería? Muchas autoridades parecen haber aceptado este hallazgo como prueba, estremecedoramente dramática, de que los brontosaurios habían sido perseguidos y caído presas de los rapaces carnívoros que les seguían la pista. Debido a que, tal como dijo una vez Richard Swann Lull, las huellas «son fósiles de seres vivos, mientras que los demás restos pertenecen a seres muertos», las pruebas de Bird han sido de gran ayuda[12]. Bird le presentó al mundo científico un breve episodio de la vida de un brontosaurio en particular; se centró en un día, día que podría haber sido testigo del final de su vida.
Evidentemente había cierto número de contradicciones. Por un lado, a los reptiles actuales se les reconocían graves limitaciones cuando debían mantener algún tipo de actividad y, a pesar de ello, los glorificados reptiles del pasado aparentemente eran capaces de las más extraordinarias hazañas. La paradoja sólo era reconocida como tal por unas pocas personas y nunca fue manifestada explícitamente, aunque implícitamente era aceptada por todos. El resultado de la lucha podría haber permanecido incierto, pero existían pocas dudas de que realmente había habido una lucha. Los carnívoros no habían sido los cobardes animales de carroña que Lambe había imaginado sino asesinos de sangre fría que aterrorizaban a sus víctimas.
Se conoce el resultado de una lucha. En agosto de 1971, la expedición paleontológica conjunta polaco-mongola al desierto del Gobi encontró los esqueletos fósiles de dos dinosaurios que, aparentemente, habían perecido mientras estaban disputando un combate a muerte. El depredador, un pequeño y ágil Velociraptor con delgadas piernas y largas manos cerradas (de aquí el nombre que significaba «ladrón rápido»), fue hallado agarrado al cráneo de un pequeño Protoceratops acorazado, abrazado inextricablemente sobre el herbívoro en una postura que sugería que ambos habían muerto en la lucha[13]. (Es un poco difícil de imaginar cómo dos combatientes podían haber muerto simultáneamente y luego haber sido conservados como congelados en la lucha). La paradoja del dinosaurio como agresor activo fue resuelta ingeniosamente por Bakker, en 1972, cuando examinó la ecología de las comunidades de dinosaurios. La relación entre depredadores y presas en las comunidades de reptiles difiere significativamente de la existente en las de los mamíferos, situación resultante de las diferentes necesidades energéticas de los dos tipos de criaturas. El lagarto vivo más grande, el dragón de Komodo (de hecho un monitor estrechamente emparentado con el mosasaurio marino del cretáceo) que sobrevive en la remota isla de Komodo al este de Bali, alcanza los 3,6 metros de longitud y un peso superior a los 450 kilogramos. Este lagarto come el equivalente al cadáver de un cerdo aproximadamente una vez al mes. Debido a que esto constituye aproximadamente la mitad de su peso, cada sesenta días habrá consumido su peso en animales de presa. Un leopardo, en cambio, necesita quemar una mayor cantidad de alimentos para satisfacer sus mayores demandas energéticas. Ingerirá su peso en alimentos en sólo diez días. Los leones consumen incluso más, su peso en tan sólo ocho días, y los perros salvajes en menos de una semana. Por lo tanto, las criaturas de sangre caliente requieren aproximadamente diez veces la cantidad de comida ingerida por un lagarto. ¿Cómo podemos aplicar estos hallazgos a las comunidades de dinosaurios de hace 70 millones de años? Una comunidad de criaturas sólo podía soportar un determinado número de depredadores para que la población se pueda mantener estable. Debido a que los carnívoros de sangre caliente matan frecuentemente, la población sólo podrá soportar a unos pocos carnívoros en relación con los inofensivos herbívoros para no alterar el equilibrio. Las comunidades de reptiles pueden tolerar un mayor número de depredadores sin que disminuya su población debido a que éstos causan un menor número de víctimas. Bakker demostró que, en las comunidades reptiles fósiles de mediados del período pérmico, existían muchos más depredadores que en las comunidades mamíferas fósiles, del oligoceno y plioceno. ¿Qué pasaba con los dinosaurios? Éstos presentaban un patrón análogo al de las relaciones presa-depredador de los mamíferos. Al igual que sucede actualmente en las llanuras de África, donde los antílopes son mucho más numerosos que los leones, en el cretáceo tardío los hadrosaurios de pico de pato y los ceratópsidos cornudos eran mucho más abundantes que los tiranosaurios contemporáneos. Los cálculos de Bakker sobre el flujo energético en las comunidades de dinosaurios, hacían que pareciese probable que la baja relación entre carnívoros y herbívoros se debiera a que éstos eran de sangre caliente y no a alguna peculiaridad en la estructura de la comunidad o incluso conservación selectiva. «El análisis de los flujos de energía indica claramente —concluyó Bakker—, que los balances de energía de los dinosaurios eran similares a los de mamíferos grandes y no a los de lagartos del tamaño de un elefante[14]». Se han dirigido críticas contra la precauciones adoptadas por Bakker, por no haber evaluado las pasiones de los recolectores de fósiles o por las irregularidades en su conservación; o, tal como destaca Alan Charig del Museo Británico, la posibilidad de discrepancias en las expectativas de vida entre los dinosaurios. Todos estos factores podían afectar drásticamente las conclusiones de Bakker. A pesar de ello, este enfoque parece ser básicamente válido, y con algunas modificaciones podría proporcionar unos excelentes resultados.
Los asesinos verdaderamente grandes de la época mesozoica tardía, los tiranosaurios y alosaurios, obviamente no eran tan manifiestos (al menos en lo concerniente a su número) como sería de esperar tratándose de lagartos monstruosos. No obstante, tal como se nos había enseñado, se trataba de bestias rapaces muy activas que con frecuencia necesitaban matar. Los Tyrannosaurus al menos, con sus grandes maxilares y pies con garras de águila, estaban capacitados para abatir a los dinosaurios con pico de pato o a los ceratópsidos. Y es muy probable que éstos fueran sus presas. Es improbable que pudieran haber cazado a los dinosaurios más pequeños y ágiles. La serie de armamentos de los tiranosaurios estaban destinados a combatir eficazmente a los herbívoros más grandes y bien protegidos como el Triceratops.
El Tyrannosaurus no estaba incapacitado para correr; probablemente, en ocasiones se veía obligado a ello para poder cazar a los galopantes Triceratops. El tiranosaurio probablemente caminaría como los patos, pero incluso los patos pueden ser rápidos cuando es necesario. Tal como suponía Barnum Brown, tanto el Tyrannosaurus como el Gorgosaurus, indudablemente eran bastante rápidos cuando mataban. Los ceratópsidos más grandes como el Chasmosaurus, que tenía el tamaño de un rinoceronte, y el Torosaurus y Triceratops, de diez toneladas de peso, ciertamente podían galopar a una velocidad notable. Al igual que los rinocerontes, los ceratópsidos estaban armados con una serie de cuernos en el cráneo (algunos de los cuales podrían haber alcanzado una longitud de 1,20 metros en vida: el núcleo óseo estaba recubierto de una funda córnea puntiaguda que se ha perdido en el fósil) y probablemente sólo se retiraban si el primer ataque no debilitaba al agresor. Bakker estimó, a partir de su estudio sobre las articulaciones de los miembros de los dinosaurios, que los grandes dinosaurios cornudos podían galopar a velocidades similares a las de los rinocerontes. «Armados con largos cuernos sobre una cabeza de gran maniobrabilidad, fuertes picos y la capacidad de galopar a velocidades de hasta 48 km/h, estos grandes ceratópsidos debieron ser los herbívoros terrestres más peligrosos que jamás hayan existido[15]». A diferencia de los tranquilos rinocerontes, los ceratópsidos tenían motivos para ser peligrosos ya que compartían su mundo con un terrible enemigo que los igualaba en fuerza.
Aquí encontramos un ejemplo grave de la endiablada hipocresía de la paleontología desde la época victoriana. Leidy y Cope no tuvieron dudas en dejar que sus dinosaurios «canguro» saltaran, y la idea de que los dinosaurios eran activos y que en ocasiones se movían con rapidez, fue arraigando de forma que, desde entonces, siempre se ha aludido a la misma. En realidad, el primer esqueleto genuino de un fósil de dinosaurio montado en los Estados Unidos fue un hadrosaurio en pleno vuelo. El problema siempre había sido lograr posturas que fueran reales. «En la mayoría de reconstrucciones —dijo Charles Beecher de la Universidad de Yale en 1902, quizá refiriéndose a los hadrosaurios de Hawkins—, generalmente se observa que el resultado concreto difiere en muchos aspectos del ideal». Las restauraciones de Hawkins se habían convertido en curiosidades históricas, así que Beecher comenzó de nuevo la rehabilitación de los dinosaurios en nuestro mundo. La dificultad era que, al no existir descendientes vivos, no se disponía de modelos para escoger la postura que debía darse al dinosaurio. Lo que se necesitaba, decía, era un esqueleto que hubiera permanecido intacto desde la muerte de la bestia, hallándose en una posición que pudiera haber adoptado en vida. Afortunadamente, los deseos de Beecher se hicieron realidad con la aparición de un magnífico espécimen del hadrosaurio Claosaunis, que había sido encontrado por los recolectores de Marsh en 1891. Sus miembros aún estaban articulados y en especial el fémur izquierdo había permanecido articulado a la pelvis. Así, el hecho de que la pierna estuviera dirigida hacia delante, como congelada a media zancada, planteó la pregunta sobre qué postura se le debía dar al animal. Beecher modeló el hadrosaurio como si estuviese corriendo, basándose en las posturas de los animales existentes en movimiento, y prestando especial atención a las fotografías del lagarto vivo Chlamydosaurus que, ocasionalmente, en trayectos cortos, corre sobre sus patas traseras.
Se ha intentado (dijo Beecher acerca de la restauración acabada del Claosaurus corriendo) llevar a cabo la idea del movimiento rápido y hacer que todas las partes del esqueleto contribuyan a la perfección y realismo del efecto general. Para lograr este objetivo debe existir un adecuado equilibrio y un balanceo verdadero del animal vivo.
Por lo tanto, se pretende que este enorme espécimen, tal como está montado ahora, transmita al observador la impresión de precipitación de una bestia mesozoica. La cabeza está levantada y girada hacia fuera. Los maxilares están ligeramente separados. Los antebrazos están balanceando los vaivenes de los hombros. La extremidad posterior izquierda se halla en un extremo de la zancada y lleva todo el peso del animal. El pie derecho ha completado el paso y ha dejado el suelo preparándose para el balanceo hacia delante. La pesada y poderosa cola está levantada en el aire y doblemente curvada para equilibrar el peso y compensar el vaivén del cuerpo y de las piernas. El aspecto global es de acción y el espectador fácilmente podrá imaginarse a esta criatura con muchos de sus atributos en vida[16].
De forma similar, Barnum Brown, dos décadas más tarde, restauró el Gorgosaurus en el Museo Americano de Historia Natural como si estuviese corriendo. Esta elección del modo de andar no es sorprendente ya que, incluso echando una ojeada rápida a los miembros y articulaciones de los dinosaurios, se puede observar que muchos dinosaurios estaban construidos de forma similar a los mamíferos ungulados modernos. Las articulaciones de los miembros de los ceratópsidos, por ejemplo, eran parecidas a las de los rinocerontes, al igual que lo era el grado de balanceo del antebrazo, lo cual inevitablemente llevaba a la conclusión de que los dos hubiesen podido llevar el mismo paso.
Los dinosaurios bípedos frecuentemente eran restaurados con las piernas en plena zancada. Una vez reconocido el hecho de que estos dinosaurios «erguidos» podían moverse con bastante rapidez, los paleontólogos comenzaron a estudiar la mecánica de los miembros y las posturas más eficaces para poder correr. Actualmente, tal como hemos visto, es corriente reconstruir estos bípedos con la cola extendida y rígida como un tendón que actúa como contrapeso y con la columna vertebral horizontal, enfundada en tendones en muchos dinosaurios para evitar que ésta se hunda. Estos revestimientos eran de máxima importancia para contrarrestar los efectos de la gravedad. En el Deinonychus, un carnívoro de poco más de 2,4 metros de largo que vivía en Montana a principios del cretáceo —y sin duda el dinosaurio más extraordinario hallado en los últimos años—, estos revestimientos habían llevado a extremos sin precedentes. Las facetas de articulación de las vértebras de la cola habían crecido formando varillas, que alcanzaban los 45 centímetros de longitud y que encajaban en las vértebras formando una unidad rígida que servía de ayuda para mantener la estabilidad dinámica. La razón de por qué la bestia necesitaba un estabilizador tan inusual se hizo evidente cuando se analizaron funcionalmente los pies. John Ostrom, que descubrió el dinosaurio en 1964, hizo estas audaces afirmaciones cuando lo presentó al mundo en 1969:
El pie del Deinonychus es, posiblemente, la prueba más reveladora, desde el punto de vista anatómico, de los hábitos de los dinosaurios en el sentido de que éstos debieron de ser cualquier cosa menos «reptiles» en su conducta, en sus reacciones y en su forma de vida. Este dinosaurio tuvo que ser un animal de carrera rápida, altamente depredador, extremadamente ágil y muy activo, sensible a muchos estímulos y rápido en sus reacciones de respuesta. Esto, a su vez, indica un nivel de actividad poco corriente para un reptil y sugiere la existencia de un índice metabólico alto. Las pruebas en favor de esa teoría radican principalmente, aunque no de modo exclusivo, en el pie[17]».
¿Qué es lo que asombraba tanto en los pies? El Deinonychus era forzosamente un bípedo ya que la estructura de sus brazos y manos mostraban claramente que el animal no podía, en ningún caso, andar sobre cuatro patas. El pie del animal se distinguía por presentar únicamente dos dedos en contacto con el suelo. Existía un tercer dedo más corto que, sin embargo, había sufrido algunas modificaciones, convirtiéndose en una estructura ofensiva que contenía una garra letal de 12 centímetros de largo en forma de hoz curvada. Este dedo se había especializado, exclusivamente, como arma extraordinaria cuya función inequívocamente era la de cortar y desgarrar.
Sin embargo, para poder matar y desgarrar a sus presas con esta garra, debía de tener un sentido del equilibrio sin par, debido a que el animal debía mantenerse de pie sobre una pata durante el proceso al mismo tiempo que luchaba con la presa. El hecho de mantener precariamente el equilibrio sobre un pie probablemente sólo se podía conseguir si la bestia disponía de una cola rígida, que actuara como una vara de balanceo incorporada, similar a la de los funámbulos. La presa debía ser mantenida alejada del cuerpo a fin de poder ser matada a golpes propinados por las patas traseras. Por esta razón, los brazos eran largos y desgarbados y las manos estaban mejor adaptadas que en ningún otro dinosaurio para agarrar y sujetar objetos. Esto le permitía al Deinonychus agarrar a la desgraciada víctima y mantenerla a distancia mientras las garras de sus pies la descuartizaban. Debido a que las cicatrices en las vértebras de este feroz asesino eran exactamente iguales a las de las grandes aves no voladoras como el emú y el avestruz, Ostrom concluyó que la columna vertebral del Deinonychus debía de haberse mantenido en una posición rígidamente horizontal mediante ligamentos. Las largas piernas revelaban al Deinonychus como veloz depredador, que fácilmente era capaz de dar alcance a sus víctimas. Esta espectacular y pequeña criatura, que se diferenciaba bastante de todas las encontradas hasta el momento (de hecho es tan diferente que Osbom no está seguro acerca de cuáles son las afinidades), apareció en un momento oportuno. En una década anterior probablemente hubiese sido visto de forma bastante distinta, y se hubiese omitido totalmente la importancia funcional de sus características más notables. Coincidiendo su aparición con la «revolución» en nuestro pensamiento, el Deinonychus encarna todo aquello que claramente no asociamos con los reptiles en los dinosaurios. Nos proporciona, declaró Ostrom al presentar a la bestia, una visión completamente nueva de las sorprendentemente sofisticadas capacidades de algunos de los dinosaurios carnívoros depredadores.
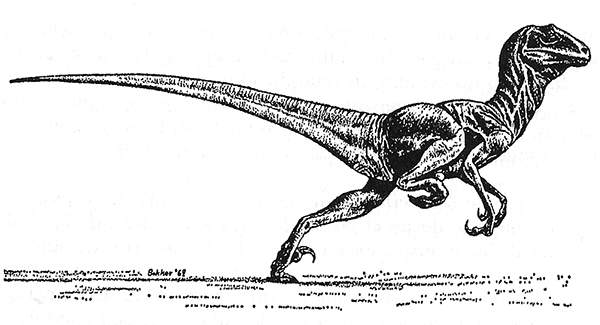
d. El Deinonychus, quizás el dinosaurio más extraordinario descubierto en los últimos años. Al igual que otros dromeosáuridos era sorprendentemente sofisticado. Reconstrucción realizada por Robert T. Bakker.
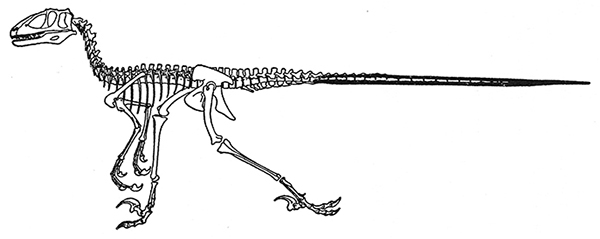
e. Deinonychus. Obsérvense las peculiares varas óseas a lo largo de toda la cola que hacían que ésta fuera rígida.
En 1970, Galton atacó de forma similar las reconstrucciones antiguas de los herbívoros bípedos, en las cuales la criatura corría con el cuerpo inclinado en un ángulo de 45 grados, con una cola fláccida arrastrando por el suelo (lo cual no solamente habría retardado considerablemente a la bestia sino que también habría causado lesiones innecesarias en la cola). En lugar de ello, también reconstruyó a estos bípedos inclinados hacia delante; esta nueva reconstrucción del hadrosaurio Anatosaurus, de cabeza plana «que corre de prisa», guarda poca relación con los intentos anteriores pero, desde luego, transmite la impresión de velocidad que anteriormente siempre faltaba[18].
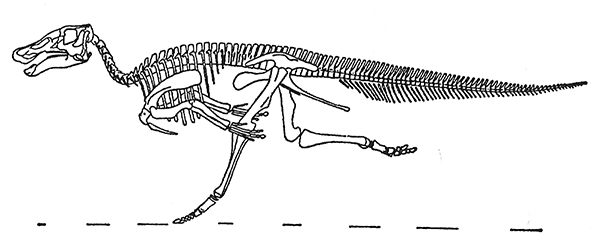
f. El Anatosaurus de pico de pato de Peter Galton «corriendo».
Sin embargo, hasta finales de la década de 1960, estas criaturas fueron consideradas básicamente como reptiles. Por muy ágil que fuera su aspecto, en vida estaban seriamente limitadas por una falta de un suministro adecuado de energía. Las apariencias naturalmente decepcionaron a muchos paleontólogos que se vieron obligados a atribuir una función a aquellas piernas largas y colocarlas de forma natural para poder escapar rápidamente cuando acechara algún peligro. Incluso a temperaturas óptimas (el momento de mayor producción de energía), los reptiles sólo pueden reunir una décima o vigésima parte de la energía de la que disponen los mamíferos de peso similar. Por lo tanto, un reptil que pese 900 gramos puede alcanzar una velocidad de 1609 km/hora con la energía disponible para ello mientras que un mamífero puede alcanzar los 16,09 km/hora o más. El Tyrannosaurus gigante quizá podía alcanzar los 4,8 o 6,4 km/hora si era ectotérmico y el Anatosaurus, incluso corriendo, no alcanzaría una velocidad mucho mayor[19].
Las criaturas más grandes y de complexión más fuerte, no muestran tan claramente lo absurdo de esta situación como lo hacen los imitadores de las aves no voladoras, con miembros largos y cuellos alargados, entre los dinosaurios, los celurosaurios. Era un hecho universalmente aceptado que los celurosaurios realmente eran criaturas muy ágiles. Colbert escribió sobre el Ornithomimus, en 1962, que «los miembros traseros son largos y están óptimamente adaptados para correr a gran velocidad[20]». El Ornithomimus, el «imitador de las aves», poseía muchas de las características que actualmente asociamos con las grandes aves no voladoras. Presentaba unos huesos finos y huecos, un cuello largo y un cráneo pequeño con un pico revestido por una vaina córnea en vida y piernas muy largas. En el imitador de los avestruces, el Struthiomimus, se habían perdido todos los dientes, lo cual aún aumentaba más su parecido con el avestruz. Al igual que los avestruces, el Struthiomimus tenía la espalda acortada, debido al acortamiento de la distancia entre la espalda y el cinturón pélvico, y los metatarsianos de los pies se habían alargado para proporcionarle un segmento funcional adicional a la pierna, permitiéndole correr sobre tres dedos (muchas criaturas veloces han sufrido estas modificaciones para incrementar la zancada). Únicamente la larga cola, los largos brazos y las manos con tres dedos hacían que este parecido superficial entre el Struthiominus y el avestruz fuera incompleto.
El Struthiomimus medía 3,6 metros de largo desde el pico hasta la punta de la cola y 2,1 metros de altura, siendo un poco más grande que el avestruz. Debido a que tenía un pico córneo sin dientes, tenemos motivos para pensar que, aunque técnicamente perteneciera al grupo de los terópodos carnívoros, la bestia se había pasado a una dieta herbívora. Los largos y delicados dedos probablemente servían para coger frutos de los árboles o para doblar las ramas hacia su pico córneo[21]. Cuando los estrudiomímidos fueron identificados por primera vez por Osborn en 1916, William K. Gregory, del Museo Americano, sugirió que las fuertes garras de los dedos, parecidas a las de los osos, posiblemente habían servido para descascarar los frutos o para coger los frutos de las palmeras, cicádidas e higueras[22]. De esta forma, los débiles maxilares desdentados podrían hacer frente a la blanda fruta carnosa. Gerhard Heilmann estaba considerablemente impresionado por las ideas de Gregory e hizo una notable reconstrucción de un grupo de estrutiomímidos, con un aspecto bastante parecido al de los avestruces (para muchas personas el parecido era incluso excesivo[23]), alimentándose de estos frutos descascarados[24]. El Struthiomimus no necesariamente debió de ser un vegetariano estricto. El avestruz es omnívoro en estado salvaje y los estrutiomímidos posiblemente también ingerían pequeños lagartos o mamíferos, que cazaban con sus manos aferradoras (ventaja que se ha perdido en el avestruz).
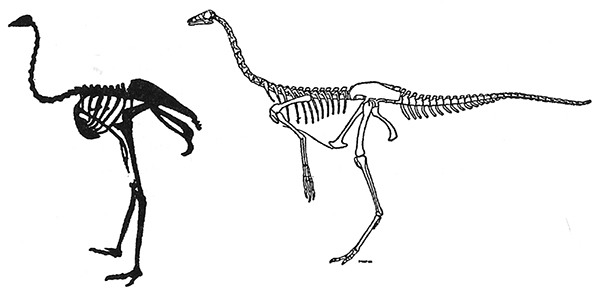
g. Una reconstrucción moderna del esqueleto del dinosaurio-avestruz Struthiomimus (derecha). Obsérvese el asombroso parecido con el avestruz (izquierda).
Las poderosas garras han sido consideradas por algunos como armas defensivas, pero es más probable que este dinosaurio con aspecto de avestruz huyera al primer indicio de peligro. Gregory no dudaba de que el Struthiomimus fuera un corredor veloz cuando estaba amenazado de peligro. Incluso lo invistió con un colgajo de piel que unía los brazos al cuerpo y que podía actuar como un plano aerodinámico para contrarrestar el peso del tronco, inclinado hacia delante cuando corría.

h. Restauración de Gerhard Heilmann del dinosaurio-avestruz.
El Struthiomimus, en realidad, no necesitaba esta ayuda aerodinámica. La cola medía 90 centímetros más que el tronco y era rígida (como en otros dinosaurios) en su mitad posterior para compensar el peso del tronco. La maravillosa similitud estructural entre el avestruz y el dinosaurio-avestruz proporciona una valiosa información al anatomista funcional acerca de los hábitos de este último. La convergencia de aspecto entre los animales siempre va unida a una estrecha similitud de las funciones. Las criaturas que tienen un aspecto parecido presentan hábitos similares; sólo hay que pensar en los Thylacosmilus marsupiales con dientes de sable y el «tigre». Smilodon con dientes de sable, ictiosaurios y delfines, etc. Estas criaturas, frecuentemente emparentadas, que en ocasiones han tenido orígenes completamente diferentes, finalmente han adquirido un aspecto similar debido a que han adoptado una forma de vida parecida. Por lo tanto, si el avestruz posee los huesos de las piernas largos para incrementar la zancada, y consiguientemente la velocidad, es razonable pensar que lo mismo es válido para el Struthiomimus, que también alargó su zancada para poder incrementar la velocidad. Debido a que los avestruces pueden alcanzar velocidades de hasta 80 km/hora, los dinosaurios-avestruces también podían alcanzar esta velocidad: al menos poseían la mecánica necesaria para alcanzarla. ¡Pero si sólo disponían del metabolismo de los reptiles, con la fisiología y producción de energía característica de los lagartos, los estrutiomímidos sólo podrían haber alcanzado la vertiginosa velocidad de 3 km/hora[25]! Esta situación paradójica, en la cual un dinosaurio tan manifiestamente construido para la velocidad estaba limitado al paso de un caracol, es equivalente a la construcción del «Concorde», equipándolo posteriormente con una maquinaria renovada de la firma «Sopwith Camel».
Gregory era un experto en locomoción animal. La velocidad, tal como demostró, está relacionada con la longitud de la zancada y la rapidez del paso. Para incrementar la longitud de la zancada, la mayoría de los mamíferos rápidos han incrementado la longitud de los huesos en la parte distal de la pata (región de la espinilla y del pie), y la comparación de las relaciones entre las longitudes de los huesos proximales y distales de la pata es un buen indicador de la velocidad que puede alcanzar un animal. El fémur de un elefante (muslo) es mucho más largo que su tibia (espinilla); el elefante es un mamífero lento y su relación T/F es de sólo 0,60. En los caballos de carreras, las tibias se han alargado considerablemente y la relación se ha incrementado a 0,92. En los velocistas realmente rápidos como las gacelas, la tibia es incluso más larga que el fémur y la zancada se acerca a su límite máximo: aquí la relación es de 1,25[26]. Los colosales brontosaurios tenían una relación T/F casi idéntica a la de los elefantes modernos; los macizos miembros y la estructura de las articulaciones de los miembros en los dos tipos de criaturas, muy separadas en el tiempo, eran asombrosamente similares y estaban diseñadas para soportar un peso aplastante. Los enormes dinosaurios también poseían unos miembros mucho más rectos que los de sus contemporáneos más ágiles y la capacidad de flexión de las rodillas y de los codos era menor, lo cual también sucede en los miembros semejantes a columnas de los elefantes. Esto hacía que los dinosaurios saurópodos caminaran muy despacio. Por el contrario, la mayoría de los hadrosaurios presentaban una relación superior a 0,80, y algunos de estos bípedos tenían las mismas proporciones que los caballos de carreras y, por lo tanto, está justificado concluir que se trataba de corredores veloces. Los dinosaurios-avestruz, tal como era de esperar, tenían una tibia que superaba en longitud al fémur, con una relación de 1,12 y, por consiguiente, la criatura probablemente podría haber competido con los mamíferos más rápidos. El singular Ornithomimus, con una relación espinilla-muslo de 1,24, era una verdadera gacela en el mundo de los dinosaurios[27].
El estudio de Dale Russell del esqueleto y de los músculos de las patas de los dinosaurios-avestruces canadienses del cretáceo tardío, y su comparación funcional con los estrutiomímidos y avestruces, han proporcionado recientemente una prueba crucial que apoya la teoría de que los dinosaurios-avestruces eran corredores veloces. Russell concluyó que, en lo concerniente a la velocidad, estos dinosaurios rara vez habían sido igualados en la historia de la vida. Al menos un miembro del grupo, el Dromiceiomimus, probablemente había sido más rápido que el avestruz[28]. Otros dinosaurios bípedos sin duda eran muy rápidos. El factor crítico que provocó que Galton sacara de una vez por todas al Hypsilophodon de los árboles fue la larga región de la espinilla. Incluso para los dinosaurios, este factor ya se había convertido en una característica indicadora de los animales corredores. El Hypsilophodon, con una relación espinilla-muslo de 1,18, debía ser capaz de alcanzar velocidades muy elevadas. Posiblemente era uno de los pocos herbívoros capaces de dejar atrás a los celorosaurios más pequeños de patas largas. El Hypsilophodon, al igual que otros dinosaurios, mantenía el cuerpo horizontal mientras corría, con la cola reforzada con tendones y alzada al aire. Al igual que otros muchos corredores, corría sobre los dedos en lugar de apoyar las plantas de los pies[29].
Estas cifras no son criterios absolutos para evaluar la velocidad. El hecho de tener unas relaciones idénticas no necesariamente implicaba que sus propietarios pudieran alcanzar velocidades idénticas en vida, ya que hay que tener en cuenta otros muchos factores (siendo uno de éstos los ajustes que deben hacerse al comparar los corredores bípedos y cuadrúpedos). No obstante, estos índices nos proporcionan una directriz. Y lo que es aún más importante, el uso de estas cifras, obtenidas a partir del estudio de mamíferos vivos y fósiles, ilustra la forma cómo los paleontólogos están empezando a estudiar a los mamíferos y aves, analizando las adaptaciones sufridas por estos animales para poder correr, a fin de comprender mejor, las criaturas que deambularon por el planeta durante la «Era de los Reptiles».
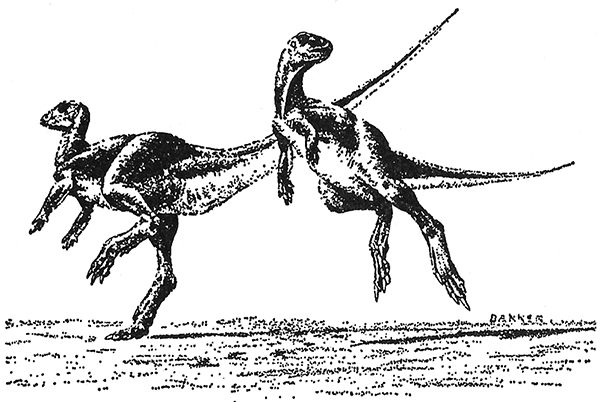
i. El Hypsilophodon, un dinosaurio pequeño y ágil, era una verdadera gacela en el mundo de los dinosaurios.