4
Todo empezó en la selva
El último ancestro común en tiempos de cambio
RELOJES MOLECULARES
Las clasificaciones del reino animal, que han permitido agruparnos juntos a una serie de especies dentro del orden de los primates, se han llevado a cabo tradicionalmente a través del estudio de las similitudes y diferencias que se observan en los órganos, músculos y esqueletos de las distintas especies vivas (lo que llamamos la anatomía comparada). Las comparaciones permiten reconstruir la historia evolutiva de esas relaciones (lo que llamamos sistemática o filogenia). Pero este tipo de comparaciones no son totalmente precisas. Por ejem plo, las dos especies de chimpancés existentes en la actualidad, el chimpancé común (Pan troglodytes) y el bonobo (Pan paniscus) son tan similares anatómicamente que no es posible saber a simple vista cuál de ellas está más cerca de los humanos. Sin embargo, a partir de la década de 1960, se pro dujo una gran revolución en la biología molecular que cambió radicalmente la forma de ver e interpretar esas relaciones. Fue entonces cuando algunos investigadores utilizaron la genética para comparar las distintas especies de grandes simios. El razonamiento era el siguiente: cuanto más próximas se encuentren dos especies, éstas compartirán un mayor número de estructuras moleculares en sus genes. Estos vínculos de proximidad informan sobre las relaciones de parentesco que se establecen entre ellas y sobre el último ancestro común que ambas compartieron (anunciado el momento anterior a su separación). Ya sabemos que los humanos compartimos el 99% de nuestro documento de identidad genético (el ADN) con los chimpancés (en realidad la cifra oscila entre el 98,8 y el 99,4%, según el procedimiento de medición que se utilice), mientras que esa similitud se hace un poco más lejana con el gorila (98%) y más aún con el orangután, con el que compartimos una semejanza del 97,6%. Los humanos estamos, por este orden, más cerca de los chimpancés y los gorilas que de los orangutanes y, por tanto, el ancestro común de estas tres primeras especies es más reciente que el ancestro común de gorilas, chimpancés y humanos con los orangutanes.
La mayor o menor diferencia genética se explica a través de las mutaciones que han experimentado nuestros respectivos genes. Si consideramos que esas mutaciones se van acumulando de forma regular a lo largo del tiempo, entonces el número de mutaciones es proporcional al tiempo que ha transcurrido desde la separación de dos ramas evolutivas. La simple contabilización de esas diferencias puede utilizarse para calcular de forma aproximada la fecha en la que una especie cualquiera comenzó su particular andadura en solitario sobre la Tierra. Esto es lo que llamamos el reloj molecular. Los especialistas en genética molecular han llegado a la conclusión de que todos los grandes simios compartimos un ancestro común que vivió hace unos 15 ó 14 m. a., momento en el que se escindió la rama que condujo a los orangutanes. De los grandes simios africanos, los primeros en separarse fueron los gorilas, hace entre 10 y 8 m. a. Finalmente, el reloj molecular indica que la separación entre las ramas que condujeron a los chimpancés y (a través de los homíninos bípedos) al Homo sapiens se produjo entre hace 7 y 5 m. a. Es en ese lejano periodo cuando el continente africano vivió una serie de acontecimientos que cambiarían el curso de las cosas. Y es exactamente en ese escenario en el que el primer acto de nuestra historia comienza.
RETRATO DE UN ANCESTRO
Reconstruir la imagen del último ancestro común entre chimpancés y humanos es una tarea relativamente sencilla. Una vez puesto a punto el reloj molecular, debemos tener en cuenta que los rasgos que definen a esa especie ancestral deben ser, como mínimo, aquellos que comparten las especies descendientes: hoy, los dos tipos de chimpancés (chimpancé común y bonobo) y los humanos. Nos parecemos tanto (genética, física y, también, culturalmente) porque somos el resultado de dos senderos que han tomado su propia dirección a partir de un punto de partida relativamente cercano. Ambos, en fin, somos hijos de una misma rama (el último ancestro común) y todos hemos pasado por una larga y paralela historia de cambios que nos han llevado a ser lo que hoy somos.
Viajemos ahora al continente africano de hace unos 8 m. a. En aquel entonces, todo el cinturón ecuatorial, desde el Golfo de Guinea (al oeste, bañado por el océano Atlántico) hasta el Cuerno de África (ya en el océano Índico, al este), estaba cubierto aún por una selva húmeda, formada al abrigo de un clima cálido y de abundantes lluvias. En estas condiciones propicias, el inmenso territorio ecuatorial se veía cubierto por una vegetación abigarrada, con gran densidad de árboles inmensos y todo tipo de plantas. En las zonas altas montañosas, las neblinas cubrían de forma habitual, casi fantasmagórica, el intenso verdor que estallaba por todas partes. En los márgenes tropicales, las selvas daban paso a grandes extensiones de bosques mixtos. Y en verdad, la cara de África era muy diferente a la actual. No solo los bosques tropicales, hoy muy retraídos, eran mucho más extensos, sino que hasta el desierto del Sahara (esa inmensa acumulación de arena abrasada) podía presumir de algunas pinceladas verdes. El paraíso vegetal selvático que hemos descrito era el hogar de innumerables especies de insectos, aves, reptiles y mamíferos que llenaban de vida y ajetreo todo el paisaje. Entre ellos, y confinado en las márgenes occidentales de la selva húmeda, hacía ya algún tiempo que el linaje ancestral que, con el tiempo, acabaría desembocando en los gorilas actuales había despegado hacia su particular aventura.
Pero fijemos nuestra atención en otro habitante de esa misma selva, ajeno ya a la suerte de los pregorilas. Se trata de un primate de no más de un metro de estatura y unos 40 kilogramos de peso, con un cerebro relativamente grande y desarrollado (no mucho más de 300 ó 400 cc.). Se mueve con desparpajo entre los árboles pero, en ocasiones y a ras del suelo, anda erguido. No desaprovecha los recursos que le ofrece la selva y se alimenta de frutas, bayas, insectos, pájaros y, cuando la ocasión se presenta, de los pequeños animales que captura. Normalmente vive en un grupo que puede llegar a reunir varias docenas de adultos con sus crías y que deambula por amplios territorios en busca del alimento. Forma parte de una sociedad extensa, dividida en grupos que, a su vez, se dividen en pequeños clanes familiares. En esa amplia red social es capaz de conservar sus vínculos a largo plazo: reconoce e identifica a congéneres a los que no ve desde hace tiempo. En su grupo se establecen relaciones basadas en una estructura jerárquica que es reconocida por sus miembros. Las relaciones entre machos y hembras incluyen algunos lazos especiales, pero los primeros se despreocupan del cuidado de las crías. Utiliza algunos artefactos (piedras o palos) para ayudarse en la recogida de alimentos o en alardes sociales y defensivos. Al igual que reconoce a los otros, tiene conciencia de sí mismo, comunica sentimientos o temores, imita, miente, negocia, manipula a los demás, se pone también en su lugar, coopera, se reconcilia y se alegra.
Por mucho que algunos se sorprendan, nuestros conocimientos sobre la vida social de los grandes simios (también orangutanes y gorilas) demuestran que es justo reconocer en ellos ciertas facultades intelectuales y sociales que hasta hace bien poco considerábamos exclusivas de los humanos. Esto no nos hace menos excepcionales ni hace menos fascinante la historia de la evolución humana, solo reconoce de una vez por todas la también excepcionalidad de nuestros parientes más cercanos. El mosaico de rasgos físicos, sociales y culturales enumerados forman parte del mínimo común denominador que compartimos los chimpancés y los humanos. Por tanto, muchos de ellos debían caracterizar al último antepasado que nos unía. A buen seguro, este gran simio vivió tranquilo en la inmensidad selvática durante algunos millones de años. Pero, ¿qué pasó?, ¿qué ocurrió para que se pusiera en marcha la locomotora de la escisión en el seno de la especie?
LA CONJURA DE LA TIERRA
En apariencia, nada catastrófico. Nada que pudiera perturbar a aquellos ancestros o que les alertara de la transformación que se avecinaba. Pero, de hecho, ese momento había llegado. La poderosa e imparable maquinaria de la naturaleza hacía tiempo que se había puesto en marcha silenciosa pero sin pausa y, aunque de forma imperceptible aún, acabaría transformando el mundo que les era conocido y afectando a algunos de sus todavía lejanos e insospechados descendientes.
Un poco después del origen de los primates (hace unos 50 m. a.), la Tierra había iniciado una nueva deriva hacia el deterioro del clima (en su larga historia ya había vivido otras dos de mayor magnitud). Sin embargo, a partir de hace unos 7 m. a. se produjo un nuevo pulso, más marcado, de enfriamiento global. La masa polar de la Antártida estaba en formación y, en el Ártico, los glaciares se extendían tímidamente. En realidad, la Tierra estaba entrando en una evidente esquizofrenia climática, en la que se alternaban cada vez más intensamente periodos de frío con otros más cálidos. Será el preludio, moderado aún, de la posterior y todavía lejana Edad del Hielo moderna (de la que, al menos en apariencia, nos hemos librado los humanos hace poco más de diez mil años). En las latitudes ecuatoriales africanas, muy alejadas de los polos y mucho menos condicionadas por el frío, estos vaivenes provocaban una sucesión continuada de fases húmedas y áridas que, a su vez, fueron responsables de la expansión y el retroceso de las selvas tropicales. Desde hace 7 m. a. el denso bosque húmedo ecuatorial se ha fragmentado y reducido en múltiples ocasiones. En la actualidad, por ejemplo, solo está presente en la Cuenca del río Congo y en África occidental.
La superficie de la Tierra está formada por inmensos fragmentos o placas que se desplazan lentamente y a la deriva, chocando unos con otros o separándose entre ellos. Estas colosales fuerzas son responsables del movimiento de los continentes, la formación de las cadenas montañosas (cuando dos placas chocan) o la apertura de grandes grietas (cuando se separan). Hace unos 30 m. a. se inició una gigantesca fractura en la corteza terrestre provocada por la separación entre las placas continentales africana y arábiga, justo a la altura de África oriental. El resultado es la imponente cicatriz del Gran Valle del Rift, que recorre más de 4.000 kilómetros desde Etiopía (al norte) hasta Mozambique (al sur). La formación del Gran Rift no ha finalizado aún y llegará el día en el que el Cuerno de África se desgaje del resto del continente. La fractura ha provocado la elevación de altas mesetas en los bordes del profundo valle y la intensa actividad geológica ha propiciado también la formación de una larga cadena de volcanes que, con sus violentas sacudidas, han expulsado una gran cantidad de rocas. Como consecuencia, se ha ido modelando una barrera que, precisamente por su elevada altitud, ha conseguido alterar la circulación de las masas de aire y bloquear la llegada de las lluvias procedentes del oeste.
Parecía, por tanto, que hace unos 7 m. a. el destino o el azar se habían conjurado para poner a prueba a nuestro protagonista. El muro del Rift no dejaba pasar las ansiadas lluvias al este del continente y, por si fuera poco, el enfriamiento progresivo de la Tierra estaba permitiendo que la di ferencia entre los periodos húmedos y secos fuera cada vez más acusada. El resultado: la selva ecuato
Una inmensa cicatriz. El valle del Gran Rift recorre África oriental de norte a sur, dividido en dos ramas, el rift occidental y el oriental. Es el resultado de la separación de las placas africana y arábiga que, en un lejano futuro, acabarán desgajando este trozo del resto del continente. rial acabaría volatilizándose de África oriental y su lugar sería ocupado por un bosque más aclarado que, solo andando el tiempo, daría paso a una sabana abierta. Y, acostumbrado al cálido resguardo de la selva ecuatorial, la sabana no parecía ser lugar para nuestro importunado ancestro.

AL ESTE DEL EDÉN
Sabemos que aquel ancestro común vivía en el cinturón selvático que un día, sin solución de continuidad, conectaba la franja ecuatorial africana de Este a Oeste. Esta estabilidad ecológica permitió también su equilibrio biológico durante muchos milenios. Sin embargo, suponemos que la barrera del Rift acabó fracturando la población inicial en dos. Los grupos que habían tenido la fortuna de encontrarse en su zona oeste siguieron gozando de las ventajas y el sosiego que les proporcionaba aquel edén selvático al que estaban acostumbrados y que seguía siendo regado generosamente por las corrientes marinas atlánticas. Serían los ancestros de los chimpancés. Aquellos a los que, por desgracia, la escisión les había encontrado en su zona oriental no corrieron la misma suerte. Serían los ancestros de los primeros homíninos y del posterior género humano. Esta teoría sobre el origen de nuestro proceso de especiación (cambio de dirección desde una especie hacia otra), formulada en 1994 por el paleontólogo francés Yves Coppens, recibe el nombre de la East side story (‘la historia del lado este’, un juego de palabras que hace referencia al musical de Broadway convertido en exitosa película en 1961). A su favor se encuentra el hecho, incontestable hasta el momento, de que entre la abultada colección de restos de animales fósiles y homíninos recuperada en los ricos yacimientos de África oriental no se ha documentado ni un solo resto fósil perteneciente a los antepasados directos de los gorilas y los chimpancés (que, por otra parte, y de forma un tanto extraña, tampoco se conocen en otros sitios). Además, la distribución actual de gorilas y chimpancés se sitúa en las regiones occidentales y centrales de África ecuatorial y no sobrepasa la frontera del Rift.
Aunque la East side story ha sido desafiada en los últimos años por algunos hallazgos, ésta se alinea con algunos de los mecanismos más vigorosos que operan en la evolución de las especies: el aislamiento de poblaciones y los cambios en el hábitat. Todos los seres vivos, incluidos los minúsculos organismos unicelulares, dependen en gran medida del medio ambiente en el que se desarrollan. Cuando se produce la transformación de su ecosistema solamente tienen dos salidas posibles: o la extinción o la adaptación a los nuevos retos (mediante la migración a zonas más parecidas a su ecosistema original o, de forma más drástica, mediante la especiación). Si, además, el reto ecológico se ve subrayado por el aislamiento reproductivo de la población, entonces la solución más drástica puede verse favorecida. La fuerza de este proceso también acabó cazando a los ancestros de los chimpancés, por ejemplo. La selva húmeda en la que quedaron inicialmente confinados tampoco fue inmune al vaivén climático del continente ni a su fracturación y retroceso. De hecho, la distribución actual de éstos es la consecuencia directa de los cambios que ha sufrido desde entonces la selva ecuatorial en África oriental y central.
En contra de lo que se pensó en un principio, los primeros homíninos no se encontraron súbitamente en medio de la sabana abierta y seca. Ese destino estaba guardado para sus descendientes.Pero sí que se vieron envueltos en un medio que ya no era el mismo, que estaba cambiando. Vieron cómo lo que antes era una selva cerrada, ahora se convertía simplemente en un margen arbolado de la misma, menos denso, más abierto. Se encontraban en una caja sin salida: de un lado, las zonas boscosas (el recuerdo de su seguro y confortable hábitat original) estaban formadas por otro tipo de árboles y arbustos; de otro, las planicies herbáceas de la sabana avanzaban peligrosamente y acabarían ganando la partida. La sabana actual es un ecosistema abierto formado por la asociación de grandes praderas cubiertas por un gran manto de plantas herbáceas y distintos tipos de árboles y arbustos de pequeñas y medianas dimensiones. Los bosques de ribera jalonan los márgenes de los ríos y lagos mientras que, a medida que nos alejamos de las zonas húmedas, las llanuras (salpicadas aquí y allá por algunas acacias) se hacen protagonistas. Debido a la gran riqueza de hierbas y plantas gramíneas (la familia de los cereales), la sabana es un paraíso para los mamíferos herbívoros (como las gacelas o las cebras) que se alimentan del pasto y son muy abundantes. Pero, precisamente por ello, la sabana acoge a grandes y peligrosos carnívoros (leones, leopardos o hienas) que encuentran su sustento en los primeros. En la sabana, las diferencias entre los periodos húmedos y secos están muy marcadas, por lo que la distribución de los recursos animales y vegetales cambia mucho a lo largo del año: en la estación seca, los herbívoros migran hacia las reservas de agua o pastos frescos y los alimentos vegetales (tubérculos, raíces y frutos) son escasos o están muy dispersos. Para los homíninos el avance de este nuevo ecosistema supondría un gran desafío a medio y largo plazo: frente a la estabilidad de la selva (donde los alimentos son constantes y abundantes durante todo el año y la cubierta arbolada ofrece refugio), la sabana representa los peligros de los carnívoros al acecho y las grandes extensiones abiertas poco arboladas, la estacionalidad acusada, la distribución dispersa de los alimentos vegetales y del agua y, finalmente, la obligación de moverse y, así, exponerse a innumerables riesgos a merced de los dictados de la naturaleza.

La sabana africana, aquí en el cráter del Ngorongoro (Tanzania), es un complejo ecosistema formado por parches de bosques abiertos y llanuras de herbáceas en las que pastan numerosas especies de herbívoros. Pero fue un medio hostil para los homíninos debido a los peligrosos carnívoros, las drásticas diferencias estacionales y la oscilación de los alimentos.
Los investigadores suponen que la adaptación a la marcha erguida debió constituir una adaptaciónrespuesta a los cambios ecológicos que comenzaron a producirse en los márgenes cambiantes de la selva en retroceso, precisamente las zonas en las que se encontraban las poblaciones ancestrales separadas ya de la rama que conduciría a los chimpancés. Es probable que esa nueva forma de desplazamiento estuviese alentada por la búsqueda de alimentos: los grupos situados al este del edén selvático pudieron habituarse a la marcha erguida como una forma más eficaz de recoger los abundantes frutos de ciertos arbustos de mediano porte que ahora comenzarían a abundar en los nuevos bosques abiertos y de transición hacia la sabana. De esta forma no habrían tenido que combinar la marcha cuadrúpeda y bípeda al desplazarse de un arbusto a otro en busca de sus ansiados frutos y podrían recolectar muchos más al mismo tiempo. Esta hipótesis, propuesta en 1994 por el primatólogo estadounidense Kevin Hunt, se ve apoyada por las observaciones del comportamiento de los chimpancés. Nuestros parientes, aunque de forma algo torpe, suelen utilizar la marcha erguida cuando están atareados buscando alimentos. Finalmente, la marcha bípeda, perfeccionada a lo largo del tiempo, habría constituido una magnífica ventaja para los homíninos, que acabarían uniendo su destino al de la exigente sabana abierta: habría permitido liberar las extremidades superiores para la talla de la piedra, para transportar alimentos desde las zonas más peligrosas a las más seguras, para atenuar los efectos del Sol en las praderas desnudas (la locomoción bípeda permite exponer menos superficie corporal a los rayos solares que la cuadrúpeda), para disponer de una mayor visibilidad y control de los riesgos en las grandes extensiones de herbáceas, o para responder con mayor eficacia a los trastornos ocasionados por la dispersión de los alimentos.
LEJANOS CANDIDATOS
Cuando los genetistas llegaron a la conclusión de que la divergencia entre los chimpancés y los humanos se debió producir entre hace 7 y 5 m. a., solo se conocían algunos dientes y fragmentos de huesos que, mal fechados, podían acercarse a los 4 m. a. de antigüedad. Los paleoantropólogos se daban cuenta, por tanto, de que el desconocimiento de este trascendental periodo para la evolución de nuestro linaje era aplastante: ¿cómo eran los antepasados de Lucy?, ¿estábamos en condiciones de identificar con cierta seguridad las líneas más ancestrales de humanos y chimpancés?, ¿era posible que el registro fósil hubiera podido conservar retazos de ese distante momento? Pronto surgieron equipos de investigación que, husmeando en sedimentos geológicos pertenecientes a ese periodo, se embarcaron en la difícil aventura de contestar aquellas preguntas. Este reto ha sido, a la vez, una reñida carrera que ha llevado consigo animadas controversias. Apenas quince años después de que la caza diera comienzo, contamos ya con algunos candidatos a hacerse con el preciado título del homínino más antiguo. Desgraciadamente, los fósiles recuperados son tan escasos y parciales que el debate sobre su inclusión o no en la rama ancestral de los humanos es muy vibrante y está completamente abierto. Esto es así porque los investigadores, deseosos de ser los descubridores de ese primer homínino (ninguno de ellos muestra tanto entusiasmo por descubrir, por ejemplo, un ancestro fósil de los gorilas), llevan al límite su interpretación de los rasgos observados. Sin embargo, debido al dinámico proceso de especiación que suponemos se inició en torno a los 7 m. a., es posible que algunas de las características que tienden a asumirse como exclusivas de los homíninos fueran, en realidad, compartidas también con otros géneros que nada tienen que ver con el linaje humano. La escasez y fragmentación de los fósiles no son, por tanto, buenos aliados para la reconstrucción de este trascendental periodo.
La huella más antigua por ahora está constituida por un puñado de fósiles recuperados en 2001, en unos sedimentos de entre 7 y 6 m. a. de antigüedad situados en la región del Sahel, en Chad (África central). Estos restos se han adscrito al nuevo género bautizado como Sahelanthropus tchadensis (‘el hombre del Sahel chadiano’), conocido popularmente como Tumai (‘esperanza de vivir’, tal y como son denominados los niños que nacen en esta región justo antes de la estación seca). El cráneo, aunque deformado, no es mayor que el de un chimpancé (370 cc.) y tampoco podemos asegurar que fuera bípedo. Sin embargo, muestra unos colmillos menos prominentes que los de nuestro pariente. La curiosa mezcla de rasgos sirvió para que sus descubridores anunciaran con gran entusiasmo que nos encontrábamos ante el representante más antiguo de nuestra estirpe, la primera evidencia de la bifurcación que conduciría hacia los humanos. No todos los investigadores están de acuerdo con esta interpretación. Algunos creen que Tumai podría ser el mismísimo ancestro común o un fósil directo de la línea que conducirá a los chimpancés. Si este ejemplar fuera realmente un homínino, su localización geográfica (en el sector centro-occidental del continente) pondría en jaque la validez de la East side story y del papel casi exclusivo de África oriental en nuestro proceso de especiación.
Otro contendiente, de edad ligeramente más moderna (6 m.a.) y descubierto en 2000 es el Orrorin tugenensis, localizado en las colinas deTugen, en el centro de Kenya, ahora sí en África oriental. Sus descubridores sostienen que ‘el hombre original’ (eso es lo que significa Orrorin) es un homínino de pleno derecho debido a que el esmalte de sus dientes es grueso (un rasgo característico de la línea que conducirá a nuestro linaje y ausente en los chimpancés) y, sobre todo, por las características modernas del esqueleto de la pierna (que, no sin gran controversia, han sido identificados como evidencias de la marcha bípeda). Aunque la desesperante fragmentación de los fósiles plantea dudas sobre su papel en la genealogía de los homíninos, su antigüedad y características lo acercan, al igual que a Tumai, a una posición ancestral.
El género Ardipithecus (ardi, en lengua afar, significa ‘tierra’), cuyos restos comenzaron a salir a la luz en 1992 en las inhóspitas tierras del norte de Etiopía, es más moderno que los anteriores y se ha dividido en dos especies: Ardipithecus kadabba (con una edad aproximada de 5,7 m. a.) y Ardipithecus ramidus (4,5 m. a.). Teniendo en cuenta la pertinaz escasez de restos, es poco lo que se sabe del grupo más antiguo. El más moderno (ramidus) presenta una dentición algo menos primitiva que el otro y, en opinión de sus descubridores, un foramen magnum más centrado que el de los chimpancés que delataría su marcha erguida.
¿Quiénes son estos enigmáticos seres? ¿Cuál es su verdadera naturaleza? La desesperante penuria de las colecciones fósiles recogidas no ayuda mucho a establecer una conclusión firme. Todos ellos poseen una talla de un simio grande; rasgos de modernidad mezclados con otros más cercanos a los de un chimpancé; estarían habituados a la vida arbórea (los huesos de la mano curvados y brazos largos son característicos en algunos ejemplares); se alimentarían de frutos, raíces, bulbos tiernos y, quizás, pequeños animales; podrían ser bípedos; podrían ser el último ancestro común entre chimpancés y humanos o podrían ser viejos homíninos (ya encarrilados en el torrente que acabará dándonos paso). Podrían ser eso y podrían no serlo. El tiempo, la fortuna de nuevos descubrimientos y la feroz competitividad de los cazadores de fósiles se encargarán de confirmar los indicios. Lo que sí sabemos es que todos ellos se hallaban en un bosque húmedo y diverso, más o menos cerrado. Aunque hoy en día la franja del Sahel es una región árida que bordea el desierto del Sahara por el sur, en aquel entonces se trataba de un paisaje bien distinto, cubierto por el colorido verdor que proporcionaban abundantes lagos, ríos, praderas y bosques, habitados por animales bien acostumbrados a una humedad generosa (cocodrilos o hipopótamos). Tumai vivió en esos parajes. Mosaicos de esa misma naturaleza, compuestos por bosques más o menos próximos, praderas húmedas y marismas, acogieron al Orrorin y a los Ardipithecus en África oriental. Estos seres, que vivieron en un periodo oscuro pero clave para la gestación de nuestro linaje, hicieron un hogar de los profundos confines de los bosques brumosos y, todavía, relativamente densos. Tranquilos en su edén quizás desconocían que todo se transforma, también el mundo que les acogía. No tenían forma de saber que sus probables descendientes se verían embarcados en una larga odisea conducida por imparables corrientes. Es cierto, soplaban fuertes vientos de cambio.
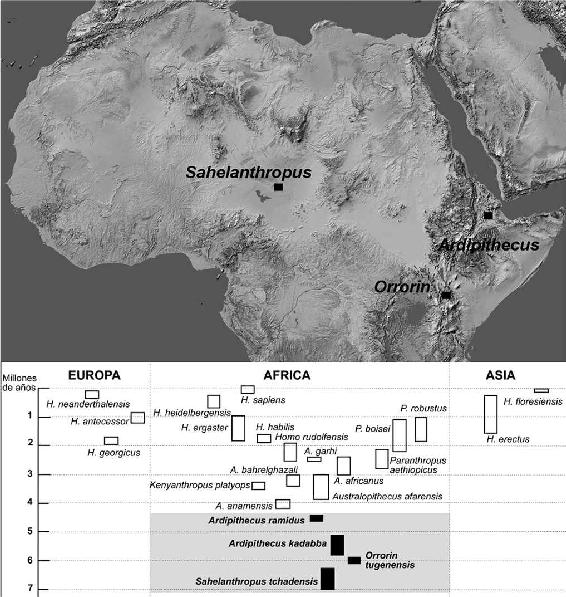
Entre hace 7 y 5 m. a. se han descubierto tres géneros distintos que pudieron pertenecer al grupo de los primeros homíninos. La escasez y fragmentación de estos fósiles plantean numerosas incógnitas sobre la naturaleza de aquellos misteriosos seres y sobre su relación con otros fósiles posteriores.

El cráneo deformado de Tumai (procedente del Chad) y reconstrucción de los parajes frecuentados por nuestros posibles ancestros. Estos seres habitaban entornos ricos en humedad y bosques mixtos. Pero una lenta cuenta atrás se había iniciado, la que impulsaba la transformación de los ecosistemas africanos.